


Sistema Endomembranar e Digestão Intracelular
📧
- Faculdade de Ciências da Universidade de Lisboa
Referência Moreira, C., (2015) Sistema Endomembranar e Digestão Intracelular, Rev. Ciência Elem., V3(4):213
DOI http://doi.org/10.24927/rce2015.213
Palavras-chave Sistema Endomembranar; Digestão Intracelular; Lisossoma; Golgi;
Resumo
A digestão é o processo de transformação das moléculas complexas em moléculas mais simples, por reações de hidrólise catalisadas por enzimas, as hidrolases. A digestão intracelular ocorre no interior da célula, por oposição à extracelular que ocorre no seu exterior.
A digestão intracelular inclui a degradação de partículas alimentares ingeridas para produzir moléculas ricas em energia – heterotrofia, e a reciclagem de estruturas celulares providenciando moléculas estruturais reutilizáveis na síntese de novos organelos – autofagia. Relacionado com os processos de digestão intracelular está o sistema endomembranar, em que existe uma relação funcional entre os seus constituintes nomeadamente o retículo endoplasmático, o complexo de Golgi e os lisossomas.
Retículo endoplasmático
O retículo endoplasmático é uma rede de membranas organizada na forma de cisterna achatadas, túbulos e vesículas esféricas, formando um sistema contínuo entre a membrana plasmática e o invólucro nuclear. O espaço interior (lúmen) está separado do citoplasma pela membrana. Existem dois tipos de retículo endoplasmático, o rugoso (RER) e o liso (REL). O retículo endoplasmático rugoso deve a sua denominação aos ribossomas associados à face externa das suas membranas que lhe conferem um aspeto rugoso quando observado ao microscópio eletrónico. O REL, pelo contrario, por não possuir ribossomas e apresenta um aspeto liso.
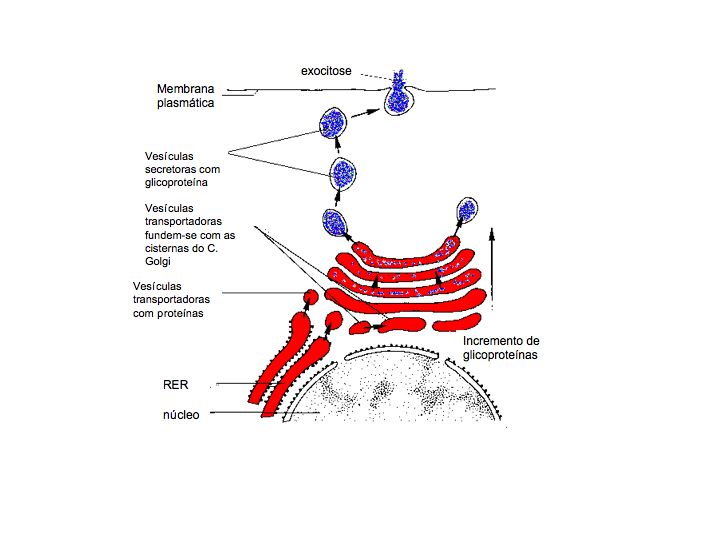
Os ribossomas associados ao RER são locais de síntese de proteínas (algumas das quais enzimáticas) com funções fora do citossol, i.e. proteínas que serão incorporadas em membranas – proteínas transmembranares, ou transportadas para outros organelos do sistema endomembranar. Estas últimas proteínas sintetizadas entram no lúmen do RER e sofrem várias modificações: conformacionais através da formação de pontes de hidrogénio e dissulfito e enrolamente da hélice adquirindo uma estrutura terciária e químicas pela adição de carbohidratos tornando-se glicoproteínas. Posteriormente, as proteínas agora designadas por proteínas secretoras, são transportadas para zonas da célula onde são necessárias ou então seguem para o complexo de Golgi, em vesículas de transporte. O RER também sintetiza ainda membrana plasmática e proteínas lisossómicas. Nas células dos mamíferos, a maioria das proteínas são transportadas para o lúmem do retículo endoplasmático à medida que são traduzidas nos ribossomas associados à membrana do RE. As proteínas cujo destino é permanecerem no citossol ou serem incorporadas no núcleo, mitoncôndrias, cloroplastos ou peroxissomas são sintetizadas nos ribossomas livres no citossol e libertadas no final do processo de tradução.
O REL é fundamental na síntese de lípidos membranares – fosfolípidos, esteroides e ácidos gordos nas células eucariotas. É também no REL que alguns carbohidratos são metabolizados e algumas substâncias tóxicas são processadas e transformadas em matérias inertes.
Complexo de Golgi
Descoberto em 1898, por Camillo Golgi nas células nervosas, e mais tarde, nos finais da década de 1950, com o auxílio da microscopia eletrónica conclui-se que existia na maioria das células eucariotas. A aparência do complexo de Golgi varia de espécie para espécie mas todos possuem o mesmo tipo de estruturas básicas, os dictiossomas. Cada dictiossoma é composto por grupos de sáculos ou cisternas membranares achatadas e empilhadas de forma regular, e por pequenas vesículas. A face convexa – região cis – virada para o RE é a face de formação das cisternas. A face côncava – face trans – é a face de maturação, onde se formam vesículas, está virada para a membrana plasmática. O complexo de Golgi está envolvido na síntese de proteoglicanos, presentes na matriz extracelular das células animais, de carbohidratos, no transporte de lípidos e na produção de lisossomas.
Algumas das proteínas sintetizadas no RER são transportadas até ao complexo de Golgi em vesículas de transporte, onde sofrem transformações permitindo que algumas proteínas se tornem funcionais e/ou algumas enzimas sejam ativadas. Após terem sofrido as várias transformações ao longo das cisternas do complexo de Golgi no sentido cis-trans, as proteínas são envolvidas em vesículas que se formam na face de maturação do complexo, as vesículas de secreção ou lisossomas.
Lisossoma
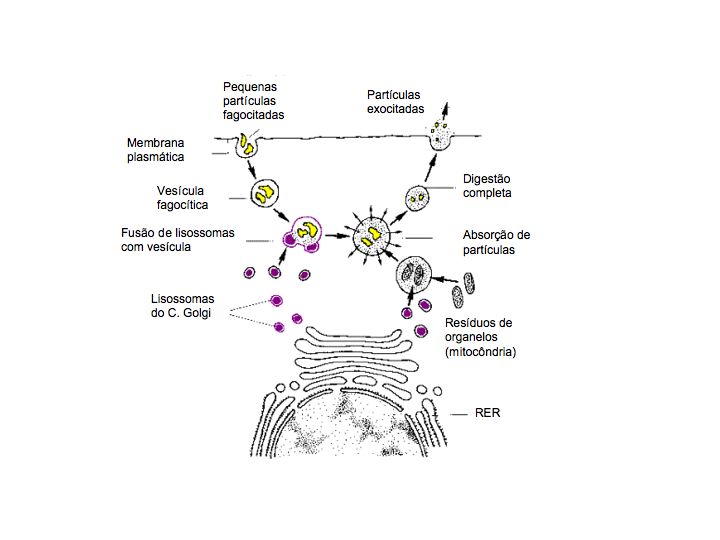
Os lisossomas são organelos esféricos membranares existentes em células animais, que contêm vários tipos de enzimas – hidrolases – que se formam na face de maturação do complexo de Golgi. Estas vesículas podem-se fundir com vacúolos ou vesículas endocíticas, formando estruturas de maiores dimensões, os vacúolos digestivos, onde ocorre a digestão – heterofagia – de muitas substâncias captadas por endocitose como proteínas. Também participam na digestão de organelos celulares – autofagia.
Este artigo já foi visualizado 14081 vezes.