


Potencial hídrico
📧
- Faculdade de Ciências e Tecnologia Universidade de Coimbra
Referência Correia, S., (2014) Potencial hídrico, Rev. Ciência Elem., V2(1):003
DOI http://doi.org/10.24927/rce2014.003
Palavras-chave Potencial; hídrico; químico;
Resumo
O conceito de potencial hídrico (ψw) é bastante importante para a compreensão das relações hídricas nas plantas e entre estas e o meio exterior (solo e atmosfera). O potencial hídrico representa o potencial químico da água, ou seja, a energia livre associada às moléculas de água. A unidade de potencial químico é o J mol-1.
Nos movimentos a curta distância, como nos sistemas osmóticos, a água desloca-se de regiões de maior potencial hídrico (maior energia livre) para regiões de potencial hídrico mais baixo (menor energia livre). O potencial hídrico de uma determinada solução é uma grandeza relativa, sendo sempre determinado em relação ao potencial hídrico da água pura medido nas condições de pressão normal e à temperatura do sistema. Assim, o ψw é calculado pela fórmula:
µw - µwº = RT ln e – RT ln eº ⇔ Δµw = RT ln e/eº (1)
em que:
- µw – potencial químico da água no sistema
- µwº – potencial químico da água pura
- R – constante dos gases
- T – Temperatura absoluta
- e – pressão de vapor da água no sistema
- eº - pressão de vapor da água pura
Se analisarmos a fórmula (1) é fácil constatar que o potencial hídrico da água pura é zero, pois se compararmos água com água o valor de pressão de vapor é igual e o ln e/eº (ln 1) é igual a zero, logo o ψw será também zero. Outra constatação que podemos retirar da fórmula é que a adição de um soluto à água, faz baixar o valor do potencial hídrico. Isso acontece porque o valor da pressão de vapor de uma solução (e) é inferior ao da água pura (eº). Deste modo, o valor e/eº será inferior a 1 e o respetivo ln um número negativo. O corolário desta situação é que a adição de um soluto à água faz baixar o valor do ψw.
Nas plantas o potencial hídrico é o resultado da soma de vários componentes, mas os mais importantes são o potencial osmótico (ψs) e o potencial de pressão ou pressão de turgescência (ψp). Assim, temos:
ψw = ψs + ψp (2)
De uma maneira geral, a adição de um soluto à água faz baixar o valor do ψw, enquanto qualquer pressão exercida sobre o sistema aumenta o valor do ψw. Ao adicionar um soluto à água, a pressão de vapor (e) terá um valor mais baixo que no caso da água (eº), em virtude de um menor número de moléculas de água se encontrar na interface líquido-vapor. Recorrendo à fórmula (1) podemos verificar que para e < eº o ln e/eº terá um valor negativo, logo o respetivo ψw será também negativo. Como é lógico, quanto maior a quantidade de soluto, menor o potencial hídrico.
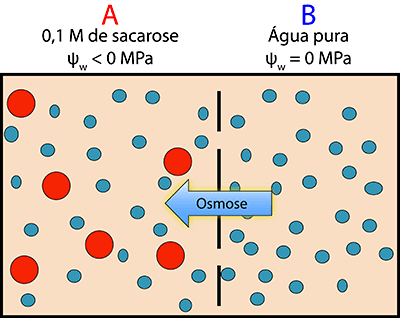
A importância do potencial hídrico e dos seus componentes pode ser ilustrada através de exemplos simples. Suponhamos que temos dois compartimentos (Figura 1) divididos por uma membrana semi-permeável (permeável ao solvente, neste caso água, e impermeável ao soluto). Consideremos ainda que o compartimento A possui uma solução 0,1M de sacarose e o compartimento B possui água pura. Se não exercermos qualquer pressão sobre o sistema, e sabendo que a água se desloca de zonas de maior potencial hídrico para zonas de menor potencial hídrico, a água deslocar-se-á predominantemente de B (compartimento com água) para A (solução de sacarose) até se atingir o equilíbrio porque o potencial hídrico da água é zero (valor mais elevado) e o potencial hídrico da solução terá um valor inferior a zero (mais baixo em comparação com a água).
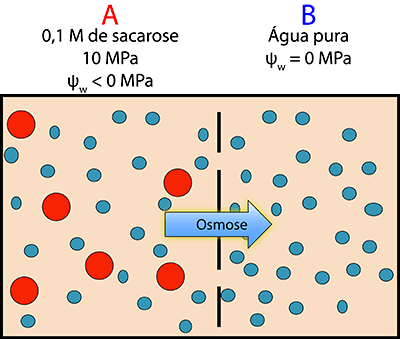
Imaginemos agora que temos dois compartimentos, como na situação anterior, em que num dos compartimentos (A) temos uma solução com um valor hipotético de ψs de -6 MPa e no outro (B) temos água (Figura 2). Se a solução em A for sujeita a uma pressão de 10 MPa como será neste caso o movimento líquido de água? De acordo com (2), o ψw no compartimento A será -6 + 10 = 4. Em B a água tem um potencial hídrico igual a zero. Deste modo, a água deslocar-se-á predominantemente de A para B pois a potencial hídrico em A é maior.
As células vegetais estão envolvidas por um meio extracelular e processos como a absorção da água ao nível das raízes, transpiração e movimentos de água entre as células e o meio extracelular são condicionados por diferenças no valor de potencial hídrico. É certo que a membrana plasmática não é uma membrana semi-permeável, pois é permeável a muitos solutos. No entanto, é uma membrana seletiva, o que significa que se comporta como uma barreira à passagem de muitos compostos.
No caso das células vegetais, e devido à existência de uma parede celular rígida a envolver o plasmalema, os movimentos de entrada e saída de água nas células são acompanhados por variações importantes no potencial de pressão, aqui designado pressão de turgescência. De facto, quando a água entra nas células, a parede impede que as células sofram um aumento de volume. Em consequência disso, desenvolve-se uma pressão que vai contribuir para alterar o valor do potencial hídrico. Assim, quando colocamos um tecido vegetal em água, esta entra nas células porque o seu potencial hídrico é maior que no interior da célula devido à existência de solutos dissolvidos. À medida que a água entra na célula aumenta o valor do potencial hídrico, atingindo-se o equilíbrio quando este for zero, tal como na água. Uma célula nestas condições diz-se túrgida. Pelo contrário, se um tecido vegetal for colocado numa solução muito concentrada (ψw muito negativo) tenderá a perder água, através do abaixamento da ψp e, consequentemente do ψw. Nestas condições as células dizem-se plasmolisadas.
Em virtude das unidades de energia serem difíceis de quantificar, os fisiologistas expressam o ψw em unidades de pressão (Pa) dividindo o potencial químico da água (J mol-1) pelo volume molal parcial da água (volume ocupado por 1 mole de água, 18 x 10-6 m3 mol-1).
Referências
- 1 Taiz, L. e Zeiger, E. (2010) Plant Physiology, 5ª Ed., Sinauer Associates, Inc
- 2 Salisbury, F. e Ross, C. (1991) Plant Physiology. 4ª Ed., Wadsworth, Belmont, CA
Este artigo já foi visualizado 14106 vezes.