


Flores, sexo e letras
📧
- CEF/ Universidade de Coimbra
Referência Canhoto, J.M., (2015) Flores, sexo e letras, Rev. Ciência Elem., V3(1):002
DOI http://doi.org/10.24927/rce2015.002
Palavras-chave flores; papoilas; humanidade;
Resumo
Tudo é mais simples do que pensas e ao mesmo tempo mais complexo do que possas imaginar.
J. W. von Goethe
As flores sempre inspiraram o génio humano. A humanidade seria certamente mais pobre sem os “lírios” de van Gogh, as “papoilas” de O’Keeffe ou as “flores” de Warhol.
No entanto, para as plantas, as flores têm funções mais prosaicas – destinam-se “apenas” a assegurar a produção de descendentes. À semelhança de muitos e diversificados organismos, as plantas também se reproduzem sexuadamente embora apresentem muitas estratégias de reprodução assexuada. Nas plantas com flor, conhecidas como angiospérmicas, as flores são os órgãos que asseguram a reprodução sexuada, sendo as angiospérmicas as únicas plantas capazes de produzir este tipo de estruturas. No entanto, a importância das flores vai muito para além da sua função biológica. O aparecimento da flor em termos evolutivos implicou o surgimento de outras estruturas, como o fruto, onde as sementes se desenvolvem e permanecem protegidas. Para além disso, em muitas espécies, o fruto auxilia a dispersão das sementes, permitindo a colonização de novos habitats. Dados recentes de biologia molecular sugerem que as angiospérmicas possam ter divergido dos seus parentes mais próximos, as gimnospérmicas, há cerca de 350 milhões de anos. No entanto, foi no Cretácico que ocorreu a sua rápida diversificação, estimando-se que há 90 milhões de anos, a maioria das cerca de 450 famílias que hoje são reconhecidas, já existiriam. Esta rápida (em termos evolutivos) radiação das angiospérmicas intrigou bastante Charles Darwin, que na sua correspondência com outros autores lhe chamou um “abominável mistério”. Estima-se que o número de espécies de angiospérmicas ronde os 350.000, o que mostra o enorme sucesso evolutivo deste grupo de plantas. A diversidade de espécies reflete-se também na diversidade floral, uma vez que cada espécie apresenta um tipo particular de flor. Além disso, em espécies que são muito utilizadas como ornamentais, como sejam as tulipas, as orquídeas ou os hibiscos, existem milhares de variedades, cada uma com um padrão de coloração diferente.

Uma flor (figura 1) completa possui quatro órgãos designados por sépalas, pétalas, estames e carpelos. Os dois primeiros não têm um envolvimento direto na reprodução, mas são muito importantes na proteção da flor (as sépalas) e na atração (pétalas) de agentes polinizadores (e.g., insetos, aves). A diversidade floral está, na maior parte dos casos, associada com modificações na forma, número e padrões de coloração destes órgãos, em particular das pétalas. Os estames e os carpelos são os órgãos reprodutores. Os primeiros são os órgãos masculinos, onde são produzidos os grãos de pólen, responsáveis pela formação e transporte dos gâmetas masculinos até ao órgão reprodutor feminino, o carpelo. Este é vulgarmente uma estrutura em forma de garrafa, com uma zona superior designada estigma onde os grãos de pólen são depositados, um canal mais ou menos comprido chamado estilete e uma zona dilatada, o ovário, onde se localiza um ou mais óvulos. No interior dos óvulos desenvolve-se um saco embrionário onde está localizado o gâmeta feminino (oosfera).
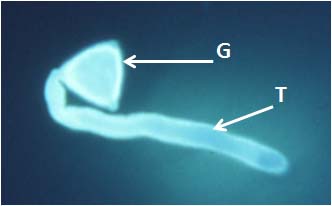
Uma vez no estigma, os grãos de pólen germinam (figura 2), ou seja, forma-se um prolongamento (tubo polínico) da célula vegetativa que vai transportar os dois gâmetas masculinos, resultantes da divisão da célula generativa, até ao saco embrionário. Para que tal aconteça, o tubo polínico tem que atravessar o estilete, que em algumas plantas pode ser longo e apresentar alguns centímetros de comprimento. A fecundação nas angiospérmicas é também diferente de outras espécies, nomeadamente das gimnospérmicas, uma vez que estão envolvidos dois gâmetas masculinos, num processo vulgarmente conhecido como dupla fecundação. Um dos gâmetas funde com a oosfera e dá origem ao zigoto, que através de um processo de desenvolvimento formará o embrião maduro e, após germinação, a nova planta. O outro gâmeta masculino funde com a célula central do caso embrionário e forma o endosperma, um tecido de reserva triploide onde se acumulam substâncias de reserva importantes para desenvolvimento da nova planta após a germinação. Por incrível que possa parecer, a humanidade depende deste processo para se alimentar. De facto, a base da alimentação mundial é o endosperma produzido por três tipos de plantas: trigo, milho e arroz. É também importante notar que o mercado da venda de plantas ornamentais representa anualmente muitos milhões de euros, existindo países, como a Holanda, onde a sua relevância é enorme. Curiosamente, neste país, no século XVII, verificou-se um interessante acontecimento que levou à ruína de milhares de pessoas e a uma importante crise económica e social. Nessa altura era moda colecionar tulipas e pagar preços muito elevados por variedades raras. Como acontece em mercados especulativos, em determinada altura, o valor das plantas caiu bruscamente, deixando muita gente na miséria. Afinal, parece que a história sempre se repete…
O resultado da dupla fecundação é dramático em termos do futuro desenvolvimento dos órgãos florais. A fecundação desencadeia a morte de todos os órgãos florais, à exceção do ovário. Este, através de uma série de modificações, origina o fruto que, à semelhança das flores, apresenta uma enorme diversidade de formas, cores e tamanhos, em função das diferentes espécies. No interior do ovário, os óvulos fecundados, sofrem também modificações profundas e originam a semente, constituída pelo tegumento (testa), o embrião e o endosperma.
Ao contrário dos mamíferos, em que os órgãos sexuais são formados durante o desenvolvimento embrionário, as plantas não possuem órgãos reprodutores durante a embriogénese e, estes, apenas se formam em plantas adultas, sendo por conseguinte resultantes do desenvolvimento pós-embrionário. Todos estamos familiarizados com o aparecimento, em determinadas alturas do ano, de flores em algumas espécies que nos são familiares. Assim, em aneiro, florescem as magnólias e as camélias, em fevereiro é comum ver surgir as flores das acácias e, na primavera, um grande número de espécies, entra em floração. Estes processos são cíclicos e para cada espécie repetem-se em alturas específicas do ano. Em muitas espécies, isto acontece porque as plantas são capazes de responder a estímulos ambientais, nomeadamente o período de luz no decurso de um dia (fotoperíodo) a que se encontram sujeitas. Esse estímulo é detetado ao nível das folhas e transportado através dos tecidos condutores (floema) para os locais onde as flores se vão formar. Durante muitos anos foi discutida na comunidade científica a natureza deste estímulo, conhecido como florígeno. Atualmente sabe-se que se trata de uma proteína produzida como resposta a alterações do fotoperíodo. As variações no fotoperíodo não são o único estímulo para a floração. Outros fatores externos, como as baixas temperaturas ou fatores endógenos como os níveis hormonais ou de nutrientes são igualmente importantes.
A formação de flores ocorre quando um (ou mais) meristema caulinar sofre uma série de modificações e evolui para um meristema inflorescencial ou floral. No primeiro caso forma-se uma inflorescência constituída por várias flores. Por exemplo, no trigo ou no girassol, as flores surgem em grupos chamados capítulos, no caso do girassol, e espigas, no trigo. Noutros casos, o meristema vegetativo evolui diretamente para um meristema floral, dando origem a uma flor, como acontece nas tulipas ou nas camélias. Isto significa que o florígeno, anteriormente referido, vai exercer o seu efeito em meristemas vegetativos, modificando a sua estrutura e fazendo com que estes meristemas passem a formar estruturas florais em vez de estruturas vegetativas (folhas).
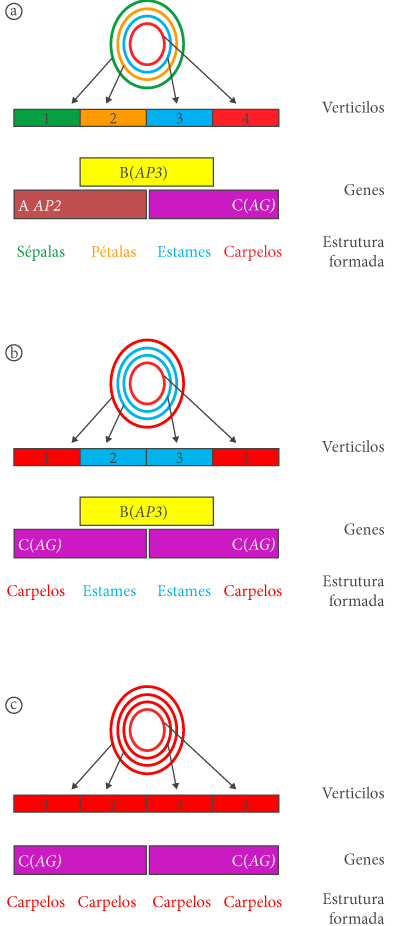
Numa flor, os órgãos florais têm uma disposição concêntrica, em anéis, que como já foi referido são quatro, sendo o anel mais externo o das sépalas, seguido pelas pétalas, estames e finalizando no anel mais interno onde se situa o carpelo. Estudos realizados inicialmente por E.S. Coen e E.M. Meyerowits e pelos seus colaboradores, e mais tarde por confirmados e aprofundados por outros autores, mostraram que existe um pequeno número de genes que interagem entre si de forma a criar este padrão de formação dos órgãos florais. Muito do que sabemos sobre o envolvimento de genes na floração resulta de estudos que têm sido realizados em plantas modelo como Arabidopsis thaliana e Antirrhinum majus (bocas-de-lobo). O modelo explicativo para a formação dos órgãos florais ficou conhecido como modelo ABC e explica a interação entre três tipos de genes que interagem para controlar a formação dos órgãos florais. Este modelo sofreu ulteriormente, modificações, com a descoberta de outros genes também envolvidos neste processo. No entanto, o modelo original é ainda em grande parte válido, pelo que será aquele aqui referido sucintamente. O modelo ABC (figura 3a) mostra que existem três genes envolvidos na formação dos órgãos florais. Como temos quatro órgãos florais e três genes, isto significa que os genes interatuam. A elaboração deste modelo resultou em grande parte da utilização de mutantes de Arabidopsis que apresentam anomalias na formação dos órgãos florais. Estas mutações afetam partes importantes do desenvolvimento e também têm sido estudadas noutros organismos, como a drosófila, para perceber como determinados genes, designados homeóticos, afetam a formação de órgãos.
Em Arabidopsis, um mutante com alteração nos órgãos florais foi designado apetala2 (ap2) sendo incapaz de formar sépalas e pétalas (figura 3b). No entanto, os quatro anéis mantêm-se sendo o verticilo exterior formado por carpelos, seguindo-se dois verticilos de estames e um último anel, mais interior, com carpelos. Assim, este mutante, apresenta carpelos no local onde se deveriam formar sépalas (primeiro anel) e estames em vez de pétalas no anel dois. Os anéis 3 e 4 têm os órgãos que surgem nas flores normais: estames e carpelos. Em termos práticos, este mutante mostra que o gene APETALA2, em condições normais (não mutado), controla a formação das sépalas e das pétalas. Uma outra mutação, chamada apetala3 (ap3) ou pistillata (pi), leva à formação de flores que possuem sépalas nos dois primeiros verticilos e carpelos nos dois mais interiores (3 e 4). Estes mutantes são incapazes de formar pétalas e estames. À semelhança do caso anterior, isto significa que o gene APETALA3, em condições normais, controla a formação de pétalas e estames. Finalmente, o mutante agamous (ag), é incapaz de formar órgãos reprodutores (estames e carpelos), apresentando pétalas nos anéis 2 e 3 e sépalas nos anéis 1 e 4. Seguindo o raciocínio anterior, esta situação mostra que o gene AGAMOUS controla a formação de estames e carpelos. Tendo em conta estes dados constata-se que a formação de sépalas é condicionada pelo gene APETALA2, a formação de pétalas pelos genes APETALA2 e APETALA3, a formação de estames pelos genes APETALA3 e AGAMOUS e que AGAMOUS controla a formação dos carpelos. Com base nestes dados, no pressuposto que os genes APETALA2 e AGAMOUS são genes cuja atividade é mutuamente exclusiva (a atividade APETALA2 impede a de AGAMOUS e vice-versa) e que a mutação num destes genes leva a que o outro expanda a sua atividade para zonas do meristema controladas pelo gene mutado, foi proposto o modelo ABC representado na figura 3. Neste modelo, a função (atividade) atribuída a cada um dos genes referidos é designada por uma letra: A para o gene APETALA2, B para o gene APETALA3 e C para o gene AGAMOUS. Imaginemos agora que temos uma dupla mutação (figura 3c) numa planta de Arabidopsis que afeta simultaneamente os genes APETALA2 e APETALA3. De acordo com o modelo, estes mutantes duplos seriam incapazes de formar qualquer órgão floral, à exceção de carpelos. De facto, a obtenção destes mutantes mostrou que as flores destas linhas de Arabidopsis possuem carpelos em todos os anéis. O leitor pode familiarizar-se com o modelo testando outras hipóteses possíveis. E no caso de um triplo mutante, ou seja, em que todos os genes referidos estejam mutados? Neste caso, verificou-se uma situação curiosa, pois todos os órgãos florais destes mutantes são… folhas. Esta situação é particularmente interessante porque o conhecido escritor e filósofo J. Wolfgand von Goethe tinha proposto (A Metamorfose das Plantas, 1790) que os órgãos florais são modificações das folhas.
Cerca de 200 anos mais tarde os dados da biologia molecular comprovaram as suposições de Goethe, o que mostra o génio do autor e como a biologia molecular é importante para compreendermos mecanismos biológicos complexos.
Jorge M. Canhoto
Departamento de Ciências da Vida
Faculdade de Ciências e Tecnologia da Universidade de Coimbra
Referências
- 1 Chase MW 2001. Angiosperms. In: eLS. John Wiley & Sons Ltd, Chichester. http://www.els.net doi:10.1038/npg.els.0003682.
- 2 Coen E 2001. Goethe and the ABC model of flower development. C.R.Acad. Sci. Paris/Life Sciences 324:523-530
- 3 Coen RS & Meyerowitz EM 1991. The war of the worls: genetic interactions controlling plant development. Nature 353:31-35.
- 4 Crepet WL 2014. Advances in flowering plant evolution. In: eLS. John Wiley & Sons Ltd, Chichester. http://www.els.net doi: 10.1002/9780470015902.q0023964.
- 5 Friedman WE 2009. The meaning of Darwin’s “abominable mystery”. Amer. J. Bot. 96:5-21.
- 6 Taiz L, Zeiger, Møller IM & Murphy H 2014. Plant Physiology and Development, 6th ed. Sinauer Associates.
Este artigo já foi visualizado 6718 vezes.