


Tecido muscular
📧
- Faculdade de Ciências da Universidade de Lisboa
Referência Moreira, C., (2015) Tecido muscular, Rev. Ciência Elem., V3(1):010
DOI http://doi.org/10.24927/rce2015.010
Palavras-chave células; tecidos; musculares;
Resumo
As células dos tecidos musculares são de origem mesodérmica. São alongadas e finas, designando-se por fibras musculares. As células são altamente especializadas, não só ao nível dos organelos mas também pela existência de filamentos, as microfibrilhas, que resultam da organização de proteínas, miosina e actina, no citoplasma, que conferem contractilidade às fibras musculares.
Os tecidos musculares (Figura 1) podem ser classificados pela sua estrutura e função em:
- tecido muscular liso;
- tecido muscular estriado: esquelético e cardíaco.
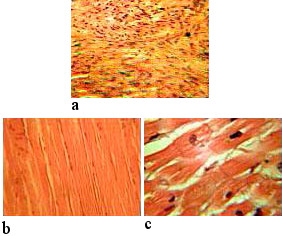
Tecido muscular liso
O tecido muscular liso é responsável pelas forças contrácteis da maioria dos nossos órgãos internos (por exemplo, vasos
sanguíneos, intestino, bexiga ou útero), que estão sob o controlo do sistema nervoso autónomo.
Do ponto vista estrutural é o tecido muscular com as células mais simples. As células são fusiformes, com um só núcleo em posição central, unidas entre si formando feixes. A sua estrutura difere bastante da dos músculos esqueléticos. A designação de tecido liso vem do facto de não ser visível estriação transversal como nos outros tecidos musculares, devido à disposição irregular das proteínas contrácteis (miosina e actina) no citoplasma.
Devem-se salientar duas características importantes no tecido muscular liso. No caso do intestino, por exemplo, as células estão dispostas em camadas e contactam entre si eletricamente através das “gap junctions”, o que permite que um potencial de ação gerado numa célula do tecido muscular liso se propague a todas as células da camada.
A outra característica das células do músculo liso é que quando as suas membranas se encontram em potencial de repouso são suscetíveis ao relaxamento. Quando as paredes do intestino relaxam e esticam num determinado local, as membranas que relaxam sofrem uma despolarização, que atingindo o limiar de potencial de ação produz um potencial de ação na membrana e as células contraem.
Assim, quanto mais relaxado estiver o músculo maior será a contração consequente.
Os tecidos musculares lisos podem ser classificados em dois grupos:
- multiunitários – com fibras individuais densamente agregadas que não se encontram ligadas entre si. Atuam de forma independente das demais fibras e são enervadas por apenas uma terminação nervosa. Exemplo, o músculo ciliar dos olhos e músculos eretores dos pêlos;
- unitários – com fibras agregadas em lâminas ou feixes, as membranas das células contactam em múltiplos pontos – junções comunicantes – através dos quais fluem iões de uma célula para a outra, de forma que contraem em conjunto. Exemplo, músculos do trato gastrointestinal, vias biliares, ou útero.
Tecido muscular estriado esquelético
O tecido muscular estriado esquelético é responsável pelos movimentos voluntários como os da locomoção (andar, correr),
pelos movimentos associados à respiração e pelo movimento individual de órgãos como o globo ocular.
São também, por isso, chamados músculos voluntários ou tecido muscular voluntário porque está subordinado a um controlo consciente.
A disposição muito organizada dos microfilamentos de actina e miosina dá-lhe um aspeto estriado. As células do musculo esquelético são de grande dimensões, e, ao contrário das do músculo liso e do músculo estriado cardíaco, possuem vários núcleos. São células sinciciais produzidas durante a fase embrionária resultantes da fusão de várias células individuais (ou da ausência de citocinese na mitose). Na grande maioria dos músculos esqueléticos as fibras estão dispostas paralelamente, unidas entre si por tecido conjuntivo. As fibras organizam-se em feixes, também estes envolvidos por tecido conjuntivo.
Cada fibra muscular é composta por miofibrilhas – feixes de filamentos contrácteis de actina e miosina. Cada miofibrilha possui filamentos finos – microfilamentos de actina – e filamentos mais grossos – microfilamentos de miosina. Num corte transversal de uma fibra, observa-se em determinado locais apenas microfilamentos de actina e noutros apenas de miosina, mas na maioria dos casos, cada microfilamento de miosina é rodeado por seis de actina. Num corte longitudinal, observa-se a estrutura estriada do músculo. O padrão estriado das miofibrilhas deve-se à presença de repetidas unidades – os sarcómeros – que são as unidades responsáveis pela contração do músculo. Cada sarcómero é constituído por microfilamentos de miosina e actina sobrepostos, quando o músculo contrai os sarcómeros encurtam.
Hugh Huxley e Andrew Huxley propuseram um mecanismo molecular para a contração muscular, deduzindo-o a partir da observação da alteração do comprimento das bandas nos sarcómeros.
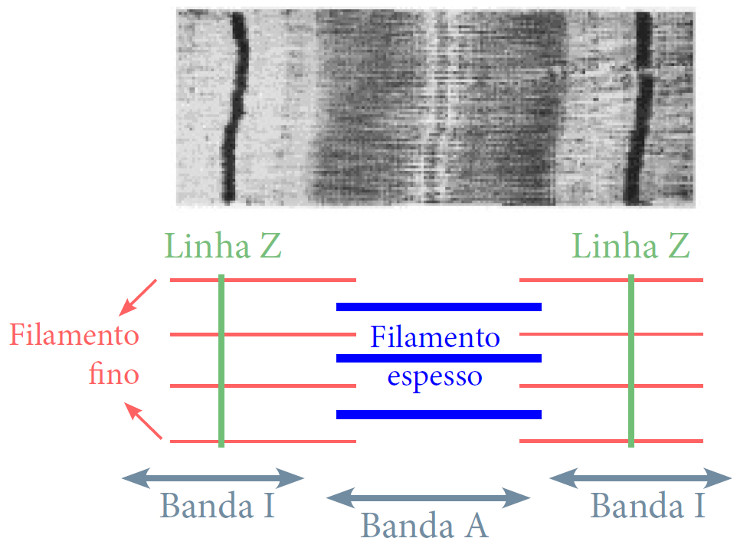
Cada sarcómero é limitado por uma linha Z (figura 2) onde estão ancoradas os microfilamentos de actina. Na zona central do sarcómero onde só existem microfilamentos de miosina, é a banda H. a banda I é a zina onde só existem filamentos de actina. A banda H e a banda I, zonas mais claras na imagem de microscópio, são regiões onde os microfilamentos de actina e miosina não se sobrepõe quando o músculo está em relaxamento. A linha mais escura na banda H é a banda M, e contém proteínas que ajudam a manter os microfilamentos de miosina organizados hexagonalmente. A banda A corresponde ao comprimento das fibras de miosina.
Quando o músculo contrai, o sarcómero encurta. As bandas H e I tornam-se mais estreitas, as linhas Z aproximam-se e deslocam-se para o centro da banda A; é como se os microfilamentos de actina deslizassem sobre os microfilamentos de miosina.
Huxley e Huxley a partir da observação desta dinâmica muscular propõem a Teoria dos Filamentos Deslizantes: segundo o modelo quando os músculos contraem os microfilamentos de actina deslizam entre os microfilamentos de miosina.
Como ocorre a contração muscular?
As moléculas de miosina possuem duas longas cadeias polipeptídicas enroladas que terminam numa cabeça globular. Os
microfilamentos de miosina são constituídos por várias moléculas de miosina dispostas paralelamente umas às outras, com as
cabeças globulares em posição lateral (ver Fig. 3). Os microfilamentos de actina são formados por duas cadeias de monómeros,
também elas enroladas entre si, e à sua volta duas cadeias de tropomiosina (proteína).
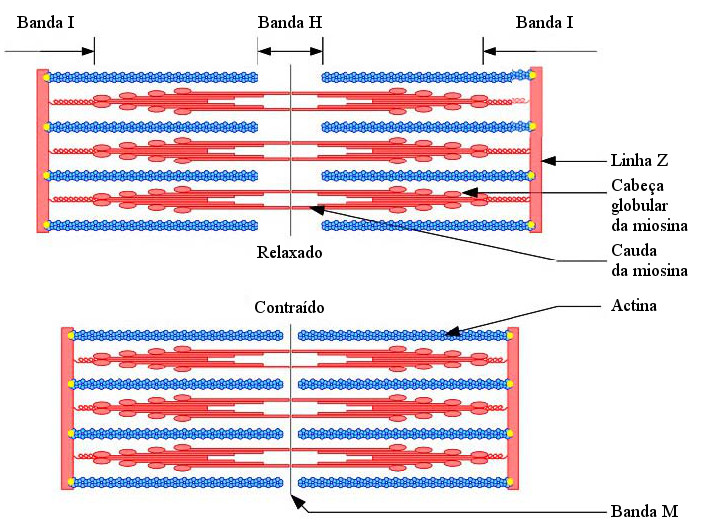
As cabeças das moléculas de miosina tem locais específicos para se ligarem à actina formando pontes entre os dois filamentos e possuem ATPase, que hidrolisa as moléculas de ATP libertando energia que é aproveitada para alterar a orientação das cabeças da miosina.
Na contração muscular, cabeça globular da miosina liga-se ao filamento de actina, alterando a sua orientação e fazendo com que os filamentos deslizem um sobre o outro. Em seguida, uma molécula de ATP liga-se à cabeça da miosina, que liberta o filamento de actina. O ATP é hidrolisado e a energia libertada é utilizada para repor a orientação original da cabeça. O ATP, curiosamente, é necessário para quebrar a ligação actina-miosina mas não para formá-las, ou seja, os músculos precisam de ATP para parar a contração (o senso comum poderia levar a pensar o contrário). Isto explica porque é que os músculos ficam rijos pouco depois dos animais morrerem, condição conhecida como rigor mortis. A morte interrompe o fornecimento de ATP armazenado nas células musculares, impedindo que as pontes entre a miosina e a actina se quebrem – o músculo não pode relaxar.
Como este processo se desencadeia em todos os sarcómeros de todas as miofibrilhas de uma fibra muscular em simultâneo, essa fibra contrai-se e todas as outras fibras do mesmo músculo.
Tecido muscular estriado cardíaco
Tem características intermédias entre o tecido muscular esquelético e o tecido muscular liso. As células são longas e
cilíndricas, como no tecido esquelético, com uma disposição das proteínas contrácteis semelhante, daí a estriação
transversal. Tal como no tecido muscular liso, as células são mononucleadas e as contrações rítmicas e involuntárias.
Algumas células do tecido muscular cardíaco podem gerar o seu próprio potencial de ação que se propaga a todo o músculo criando o ritmo cardíaco.
Além destas características partilhadas, o tecido muscular cardíaco tem características únicas: as fibras são ramificadas em Y, unindo-se longitudinalmente às células vizinhas através de estruturas designadas por discos intercalares.
Entre as células existe tecido conjuntivo que suporta uma rede capilar fundamental para o metabolismo intenso, conferindo-lhe resistência às grandes pressões a que estão sujeitas durante o bombear de sangue no coração.
Este artigo já foi visualizado 28280 vezes.