


Madeira
📧
- CEF/ Universidade de Coimbra
Referência Canhoto, J.M., (2018) Madeira, Rev. Ciência Elem., V6(4):074
DOI http://doi.org/10.24927/rce2018.074
Palavras-chave Madeira; material biológico; árvores; angiospérmicas; gimnospérmicas;
Resumo
A madeira é um material biológico produzido pelas árvores que se acumula em grandes quantidades no seu tronco. Trata-se de um tecido formado por vários tipos de células que têm em comum o facto de possuírem na parede um composto químico extremamente resistente que lhes confere rigidez e a capacidade de suportar tensões ou pressões elevadas. Os botânicos chamam à madeira lenho ou xilema secundário. A função deste tecido é conduzir a chamada seiva bruta, formada essencialmente por água, elementos minerais e alguns compostos orgânicos, desde a raiz até à parte aérea onde estes componentes vão ser utilizados no metabolismo das plantas. Para além disso, a madeira funciona também como um esqueleto, uma vez que garante a sustentação das árvores, permitindo-lhes alcançarem grandes alturas e competirem de uma forma eficaz pela luz. A madeira é utilizada pelos humanos desde tempos imemoriais, não apenas para a produção de energia, mas também na construção e no fabrico de papel. A madeira das árvores tem características distintas consoante as árvores são folhosas, como o sobreiro e o choupo ou resinosas, como os pinheiros e os teixos.
Os organismos a que chamamos árvores pertencem a dois grandes grupos de plantas, as angiospérmicas e as gimnospérmicas. Mais especificamente ainda, pode dizer-se que praticamente todas as árvores são dicotiledóneas ou coníferas. Os viveiristas e os madeireiros chamam a estes diferentes tipos de árvores folhosas e resinosas, respetivamente. As folhosas são constituídas por dicotiledóneas arbóreas e a sua madeira é, em regra, mais densa que a madeira das resinosas, que como o nome indica, são também capazes de produzir resina. A madeira destes dois grupos é diferente porque, as células que a constituem são também diferentes, existindo uma maior diversidade celular na madeira das folhosas do que na madeira das resinosas.
Do ponto de vista evolutivo, o aparecimento de um tecido especializado como o xilema, foi um aspeto importante para a conquista pelas plantas dos habitats terrestres. De facto, as plantas mais primitivas, como os musgos, não possuem tecidos especializados no transporte de nutrientes, como o xilema ou o floema. Esta característica só surge com as pteridófitas (e.g., fetos) e atinge a sua expressão máxima nas angiospérmicas e nas gimnospérmicas. À exceção das briófitas todas as outras plantas possuem células de xilema especializadas na condução da seiva bruta.
Em termos biológicos, a madeira não é mais do que o xilema secundário, um tecido complexo, constituído por células condutoras, células de suporte e células de parênquima que, no seu conjunto, conferem à madeira as suas propriedades, variáveis segundo as diferentes espécies. Do ponto de vista evolutivo, o aparecimento de um tecido especializado como o xilema, foi um aspeto importante para a conquista pelas plantas dos habitats terrestres. De facto, as plantas mais primitivas, como os musgos, não possuem tecidos especializados no transporte de células, como o xilema ou o floema. Esta característica só surge com as pteridófitas (e.g., fetos) e atinge a sua expressão máxima nas angiospérmicas e nas gimnospérmicas. À exceção das briófitas todas as outras plantas possuem células de xilema especializadas na condução da seiva bruta. No entanto, o xilema secundário, está apenas presente nas espécies que apresentam aquilo a que se chama crescimento secundário. Este tipo de desenvolvimento das plantas, ocorre não apenas em árvores e arbustos, mas também em alguns órgãos utilizados na alimentação humana, como a cenoura ou a beterraba. No entanto, o crescimento secundário que ocorre no tronco de uma árvore é completamente diferente daquele que ocorre numa cenoura e distingue-se pelo tipo de tecidos produzidos. Assim, enquanto numa árvore o crescimento secundário origina em grande parte xilema secundário, na raiz da cenoura o principal tecido produzido é o parênquima que tem essencialmente uma função de reserva, acumulando amido e pigmentos.
O crescimento secundário (em largura) das espécies arbustivas e arbóreas, deve-se à existência de dois meristemas laterais, assim designados por oposição aos meristemas primários, que se localizam na extremidade dos ramos e das raízes e que são responsáveis pelo alongamento dos caules e raízes, aquilo a que se chama crescimento primário. Esses meristemas são o câmbio vascular e o felogénio, este último também chamado câmbio súbero-felodérmico (FIGURA 1). Este último câmbio é responsável pela formação daquilo a que chamamos a casca das árvores. Por exemplo, no sobreiro, a cortiça (súber) resulta da atividade do felogénio. Dada a função protetora das células que origina, este câmbio tem uma posição mais periférica.
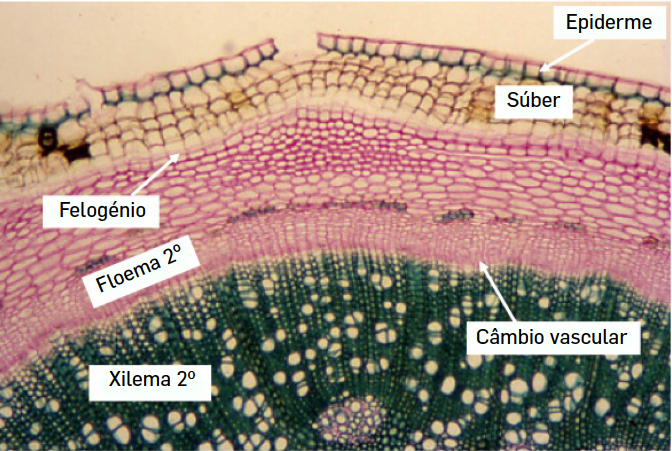
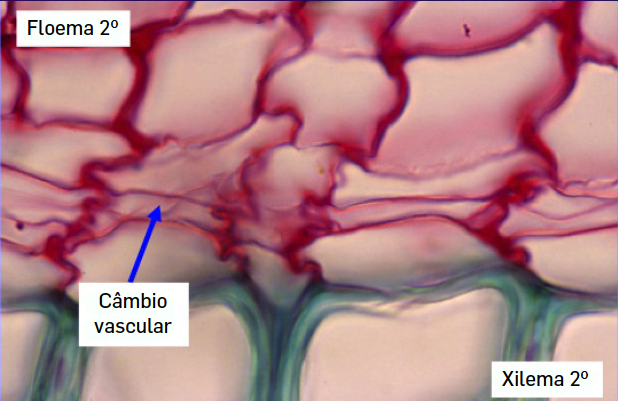
O câmbio vascular tem uma posição mais interna e produz dois tipos de tecidos especializados na condução de substâncias, floema e xilema (FIGURA 2). Por serem produzidos a partir deste meristema secundário, estes tecidos são designados por floema e xilema secundários, para os distinguir do floema e xilema primários que são produzidos a partir de um outro meristema (primário), o procâmbio. Importa referir que todas as plantas, mesmo as herbáceas, possuem xilema para a condução da seiva bruta. O que nem todas as plantas possuem é xilema secundário, pois o câmbio vascular só está presente em algumas espécies. Na realidade, o que distingue o xilema das plantas herbáceas e das plantas arbóreas é a sua origem e a quantidade, pois os tipos celulares que o constituem são os mesmos.
Se a madeira deriva do câmbio vascular e se este meristema produz dois tipos de tecidos vasculares, o floema e o xilema secundários, não deviam os troncos das árvores ter quantidades semelhantes destes dois tecidos? A resposta seria sim, se o câmbio funcionasse de forma simétrica, mas o que se passa na realidade é que o câmbio produz muito mais xilema secundário do que floema secundário. Isto quer dizer que as células derivadas do câmbio, se diferenciam muito mais em células de xilema do que em células de floema. Outra particularidade do câmbio vascular, é que ele forma um anel contínuo formado por uma única camada de células de espessura. As células que se diferenciam para fora do anel do câmbio (em direção à epiderme) vão diferenciar-se em células de floema enquanto aquelas que se diferenciam para o lado de dentro do anel irão evoluir para células de xilema. Desta forma, o xilema secundário tem uma posição mais interna que o floema secundário.
As células do xilema apresentam várias particularidades que as distinguem de outros tipos celulares e que lhe conferem propriedades específicas que permitem, por um lado, a condução da seiva bruta, mas também contribuem para a sustentação dos órgãos vegetais, uma vez que as células possuem uma parede celular rica em lenhina, um dos compostos biológicos mais resistentes que se conhecem. Quando completamente diferenciadas, as células do xilema secundário são células mortas em que o citoplasma degenerou permanecendo apenas a parede celular. Excetuam-se as células vivas de parênquima que constituem os raios medulares envolvidas no transporte lateral de nutrientes e que colocam em ligação o floema e o xilema. Células do parênquima estão também em contacto com as células condutoras do xilema, aquilo a que se chama o parênquima axial.
A parede celular está presente nas células de diferentes organismos, como acontece em algas, fungos e plantas. No entanto, a sua estrutura e composição é diferente nestes organismos. No caso particular das células vegetais, a parede assume também características bioquímicas e estruturais muito diferentes em função dos tipos celulares. Todas as células possuem uma parede primária, exterior à membrana plasmática e que é rica em celulose. As fibras de celulose estão embebidas numa matriz formada por distintos tipos de polissacarídeos que são, no seu conjunto designados por pectinas e hemiceluloses. Elementos minerais, proteínas (estruturais e enzimáticas) e compostos fenólicos estão também presentes. Algumas células formam, internamente à parede primária, uma parede secundária. Esta parede é química e estruturalmente muito diferente da parede primária, pois é rica em lenhina, que rodeia as fibras de celulose, e pode possuir diferentes camadas. Esta parede confere às células uma forte sustentação mecânica permitindo que estas possam sofrer tensões e pressões elevadas. A parede secundária é típica das células xilémicas e do esclerênquima.
Ao analisarmos as células do xilema podemos verificar que existe alguma diversidade nas células que estão presentes e que essa diversidade é notória quando comparamos o lenho de folhosas com o lenho de resinosas. Nestas últimas, o lenho é mais simples (homogéneo) e apresenta-se formado por três tipos celulares: células de parênquima, células com função de suporte (fibrotraqueídos) e células com uma função condutora (traqueídos). Deve no entanto referir-se que os traqueídos desempenham também uma função de suporte devido às suas paredes espessas. Os traqueídos e os fibritraqueídos são células normalmente orientadas segundo o eixo do caule (eixo axial) enquanto que as células de parênquima se organizem em raios (raios medulares) com uma orientação perpendicular (eixo radial) às primeiras.
Os traqueídos são células alongadas, fechadas nas extremidades, em que a seiva se desloca de uma célula para outra através de orifícios que existem na parede celular, chamados pontuações. As pontuações dos traqueídos são estruturas complexas constituídas por parede primária e secundária que permitem controlar o movimento da seiva bruta. Nas folhosas, o xilema é mais complexo (heterogéneo), pois para além dos tipos celulares já referidos, existem dois outros tipos celulares muito especializados, os elementos de vaso e as fibras. Estas últimas, tal como os fibrotraqueídos, são células com funções de suporte, muito alongadas, com uma parede muito espessa e um lúmen bastante reduzido. Os elementos de vaso são células perfuradas nas extremidades, de diâmetro muito maior que os traqueídos e que se alinham topo a topo. A seiva bruta passa de uma célula para outra através das placas de perfuração que se situam nos topos das células. Em virtude dos elementos de vaso serem células de maior diâmetro que os traqueídos e das suas extremidades estarem perfuradas, a velocidade de circulação da seiva bruta nas folhosas é maior do que a que se verifica em coníferas.
Uma particularidade da madeira é que a maneira como as células se distribuem não é uniforme, sendo visível em muitas espécies um padrão de bandas que corresponde aos anéis de crescimento (FIGURA 3).
Este padrão é típico de árvores de climas temperados, quer se trate de dicotiledóneas ou de coníferas, em que as condições ambientais são muito diferentes ao longo do ano, mas ocorre também em espécies tropicais, em zonas onde existe uma estação mais chuvosa e uma estação onde a pluviosidade é mais reduzida. No entanto, em climas tropicais, vários anéis, por vezes quase impercetíveis, podem ocorrer no mesmo ano, tornando, difícil relacionar o número de anéis com a idade das árvores. Os anéis de crescimento são um exemplo de plasticidade fenotípica e mostram como o ambiente condiciona o desenvolvimento dos organismos.
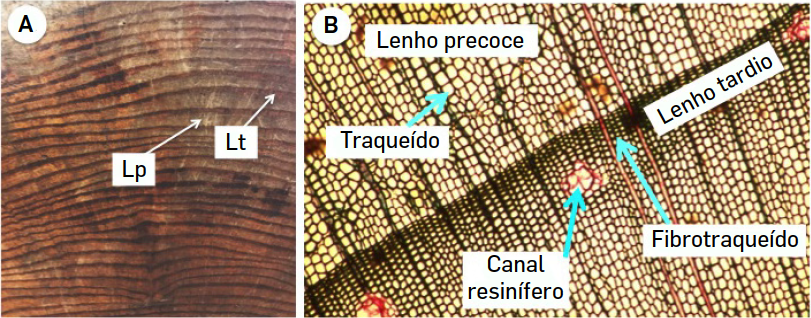
Se se reparar num anel de crescimento, verifica-se que ele é formado por duas zonas bem distintas, uma mais clara e outra mais escura. A zona mais clara corresponde ao chamado lenho precoce ou primaveril enquanto o anel mais escuro corresponde ao lenho tardio ou outonal. Estas diferenças resultam da atividade do câmbio vascular. Quando as condições ambientais são mais favoráveis ao crescimento, o câmbio produz o chamado lenho primaveril formado essencialmente por células condutoras, traqueídos nas coníferas e elementos de vaso nas dicotiledóneas. No final do verão e princípio do outono, o câmbio começa a ter uma atividade mais reduzida. Em consequência disso, o número de células formado é mais reduzido e o tipo celular mais comum são células de suporte, fibras nas dicotiledóneas e fibrotraqueídos nas coníferas. No inverno, o câmbio está inativo e não produz células. Na primavera seguinte, o câmbio retoma a sua atividade e um novo anel é formado. Num corte transversal de um tronco com muitos anos é possível verificar que os anéis formados quando a planta era mais jovem (anéis mais interiores) são mais largos, pois as taxas de crescimento em árvores jovens são mais elevadas que em árvores adultas. É também possível verificar que nem todos os anéis mais exteriores possuem a mesma espessura, fruto de condições ambientais que variam de ano para ano (disponibilidade de água, temperatura, luminosidade...). Deve ainda referir-se que numa árvore, apenas os anéis mais exteriores apresentam células funcionais, capazes de conduzir a seiva. Nos anéis mais interiores acumulam-se resinas, gomes, compostos fenólicos e muitos outros compostos nas células que preenchem o lúmen e impedem a translocação de nutrientes. Esta parte mais interna da madeira tem normalmente uma coloração mais escura e é designada por cerne ou durame (FIGURA 4). A parte mais externa, normalmente mais clara, é o borne ou alburno. Devido às suas diferentes características, estes dois tipos de madeira têm propriedades diferentes o que se reflete, por exemplo, no fabrico de mobiliário. A coloração mais escura do cerne é também muito apreciada pelos consumidores e a impregnação das células com os compostos acima referidos, associada ao seu menor teor em água, faz com que a xilema secundário desta parte da árvore seja mais resistente às condições ambientais e aos organismos que atacam a madeira. O estudo dos anéis das árvores (dendrocronologia) tem importantes aplicações práticas como sejam a datação de edifícios antigos, de pinturas e na evolução das condições climatéricas.

Numa época em que muito se fala de alterações climáticas e da origem antropogénica do dióxido de carbono atmosférico, convém salientar que as árvores, devido à madeira que produzem, são importantes sumidouros de carbono, contribuindo para mitigar o efeito de uma acumulação excessiva deste gás.
Referências
- 1 Crang, R., et al., Plant Anatomy. Springer 2018.
- 2 Cutler, D. F. et al., Plant Anatomy – An Applied Approach. Blackwell Pubishing, 2008. ISBN: 978-1-405-12679-3
- 3 Etchells, J. P. & Turner, S., Lateral meristems. In: eLS. John Wiley & Sons Ltd, Chichester, 2009.
- 4 Evert, R. F., Esau´s Plant Anatomy, 3ª ed. Wiley-Interscience, New Jersey, 2006.
- 5 Fischesser, B., Conhecer as Árvores, 3ª ed. Colecção EuroAgro, Portugal, 2005.
- 6 Hough, R. B., The Woodbook. Taschen, Hong Kong, 2002.
- 7 Myburg, A. A. et al., Xylem Structure and Function. In: eLS. John Wiley & Sons Ltd, Chichester, 2013.
Este artigo já foi visualizado 9134 vezes.