


A dualidade gradualismosaltacionismo e a escala analítica
📧 , 📧
- * CITAB/ UTAD
- ɫ CITAB/ UTAD
Referência Zambruno, J. M., Crespí, A. L., (2022) A dualidade gradualismosaltacionismo e a escala analítica, Rev. Ciência Elem., V10(3):047
DOI http://doi.org/10.24927/rce2022.047
Palavras-chave
Resumo
A análise do desenvolvimento morfogenético evidencia a relevância da escala analítica, especialmente em relação à existência de planos corporais e, portanto, à linearidade filogenética. Nesta discussão, a continuidade e descontinuidade, expressada em forma de gradualismo ou saltacionismo, ganha uma perspetiva complementar e não excludente.
A variabilidade perante o desenvolvimento e evolução
Independentemente das interrogantes intrínsecas à transição de organismos unicelulares a pluricelulares, ou da suposta eficiência da agregação celular em relação à vida unicelular, a presença de agregados não somente consolida a organização de funções. Esse esforço por agrupar e organizar funcionalidades transformou-se, ao mesmo tempo, numa corrida imparável pela diversificação de formas e comportamentos.
Neste sentido, é indiscutível que, através da união e coordenação de múltiplas células, os seres pluricelulares alcançaram vantagens inquestionáveis, especialmente em relação à diversificação das formas. Já desde uma perspetiva funcional, a multiplicidade de recursos que podem ser utilizados de forma simultânea, com a mesma ou com diferentes finalidades, é uma outra vantagem a ter presente. O aumento do tamanho inerente envolve, igualmente, uma redução do risco de depredação, ao mesmo tempo que amplia a superfície de interação com o meio. Esta última caraterística incrementa a incorporação de recursos extracelulares, bem como a possibilidade de atrapá-los e poder manipulá-los. Com efeito, e para além destas e outras caraterísticas, a diferenciação organográfica supõe uma consolidação dos mais diversos fins, bem como um elevadíssimo alargamento de opções nos processos de desenvolvimento1.
Por meio da diversidade de funcionalidades e formas, reforça-se, igualmente, a ideia da complexidade pluricelular, neste caso como acesso obrigado para a “variabilidade”. É assim que os seres pluricelulares constroem a sua diversificação morfogenética. Através desta variabilidade, são atingidos processos de divergência evolutiva, em que determinados grupos isolam-se reprodutivamente dos ancestrais. Aqui surge uma primeira questão: seria factível a existência de uma planificação desses processos de desenvolvimento?
Planos de desenvolvimento no conflito entre saltacionismo e gradualismo
A ideia de uma suposta planificação nas formas representadas pelos seres vivos começa a ganhar corpo desde a sistemática aristotélica. Esse sentido classificativo, marcado pela filosofia científica helénica, determinaria aquilo a que Darwin designaria de “planos corporais” (body plans). Esta hipótese darwinista, destinada a descrever as consequências dos processos evolutivos2, acompanharia a polémica entre a postura gradualista defendida pelo próprio Darwin3, em contraponto com o saltacionismo, mais tarde consolidado pelos mendelianos.
A polémica entre continuidade e descontinuidade poderia ser enquadrada em quatro critérios descritivos diferentes: o seu significado funcional, evolutivo, homólogo e qualitativo4. A partir destas visões do processo evolutivo, o desenvolvimento associado (expressado através da variabilidade morfogenética e ambiental) acabaria por ser exposto à compatibilidade reprodutiva entre indivíduos, de modo a gerar novas funcionalidades (como bem defendia já o próprio Cuvier)5. Esta dinâmica funcional permitiria explorar caraterísticas morfogenéticas, ambientais e comportamentais, com as que encontrar novos nichos (processos designados por key innovations)6, 7, potencialidades originadas desde a homologia funcional8, 9 ou desde as diferenças qualitativas retiradas da exploração de determinados carateres (sustentado sobre a capacidade dos indivíduos de explorar interações epigenéticas nos sistemas em que se desenvolvam)10. Todos estes mecanismos denunciam descontinuidades, mas também em todos eles é, igualmente, factível a concretização de gradualismos, com os que traçar linhas filogenéticas.
Perante um cenário como este, e tendo presente o comportamento sistémico inerente ao funcionamento (ou seja, do conjunto dos indivíduos dentro dos seus sistemas ecológicos e da sua interação com a matriz ambiental), constrangimentos das mais diversas intensidades e durações vão surgindo ao longo do espaço e tempo. Precisamente, a perspetiva funcional do sistema acaba por promover a existência de aparentes planos corporais próprios para cada grupo biológico (espécies, géneros, famílias ou, de modo mais genérico, clades). Sobre esses planos morfogenéticos, sustentam-se os processos de desenvolvimento e evolução. Desta forma, e com o propósito de descrever esses percursos em que os indivíduos respondem ao longo do tempo à heterogeneidade e dinâmicas ambientais, as escalas analíticas desempenham aqui um papel decisivo. É necessário não esquecer que as continuidades funcionais dependerão sempre das unidades escolhidas na descrição desses processos. A granulação ou descontinuidade analítica implicaria, neste sentido, a presença de supostos saltos comportamentais, que unicamente não seriam senão o resultado da escala à qual tivesse sido realizada a análise funcional. Sendo assim, perspetivas macroanalíticas proporcionam panoramas claramente particulados, em que conjuntos de planos corporais e os seus respetivos desenvolvimentos envolvem variabilidades morfogenéticas presentes no sistema ou conjunto de sistemas estudados. Por esta razão, até processos alopátricos ou parapátricos, resultantes de ruturas populacionais abruptas e alongadas, deveriam responder, igualmente, a fenómenos evolutivos contínuos e, portanto, graduais, por forma a não quebrar funcionalidades do sistema.
A origem da polémica
O problema parece estar concentrado na escala analítica e, portanto, nas implicações funcionais dessa perspetiva de caraterização. Para tentar compreender a polémica envolta entre gradualismo e saltacionismo, é imprescindível recorrer, em primeiro lugar, à origem da dúvida: o próprio Darwin. Para o autor de “A origem das espécies”, a dificuldade em explicar qualquer gradualismo entre planos corporais de uma mesma linhagem devia-se à falta de informação paleontológica11. Para entendermos esta possível explicação, é imprescindível não esquecer que o Darwinismo clássico é, antes de nada, defensor do ambiente como “mono-factor” propulsor da seleção natural12. Aqui estava a enorme dificuldade expressada pelos darwinistas clássicos, por forma a explicar as variações funcionais ao longo dos planos corporais de uma mesma linha filogenética13.
O acesso do mendelismo e dos fenómenos de mutação genética, como explicação da variabilidade molecular das cadeias polinucleotídicas, proporciona a visão saltacionista, defendida por Bateson e Goldschmid14, mais tarde consolidada por Gould e Eldredge15. A reconciliação entre darwinismo e mendelismo tenta conjugar gradualismo e saltacionismo no neodarwinismo, mais tarde reunido na Moderna Teoria Sintética da Evolução (MTE), sujeita a posteriores extensões procedentes de novos contributos para o estudo filogenético13, 14.
Nesta progressão do pensamento evolucionista, a perspetiva darwinista da variabilidade ganha um especial reforço, neste caso através da análise do “desenvolvimento”. Aquilo que poderíamos designar por origem de um novo morfogenótipo e, portanto, de uma divergência evolutiva, tem como fundamento um desenvolvimento prévio16. Sobre essa divergência, fruto de uma mutação genética ou de alterações ambientais, incidirão pressões seletivas posteriores. Precisamente neste ponto, concentra-se também a origem da discussão. Somos capazes de descrever planos corporais, proporcionando para isso processos de continuidade que estabeleçam semelhanças e, desta forma, confirma- se a existência desses planos. Mas, é nessas alterações ou aparentes descontinuidades, surgidas ao longo desse percurso, que não conseguimos determinar a natureza dessa suposta divergência.
Para melhor compreendermos o problema gerado pelo dilema gradualista/saltacionista, podemos fazer um exercício de imaginação, através do qual criemos uma hipotética situação em que uma alteração genética despoleta uma variação fenotípica determinada. Essa variação estará acompanhada por um conjunto de desenvolvimentos morfocomportamentais, que consigam dar reposta aos desafios ambientais. Em princípio, este processo de desenvolvimento não obrigaria a isolamentos reprodutivos, uma vez que esse fenómeno impediria a expansão da variabilidade genética envolvida. No caso de vir a surgir algum isolamento reprodutivo estaríamos perante um fenómeno saltacionista? Obviamente não, mas poderá vir a ser a origem de uma provável divergência dentro do mesmo plano corporal, sempre que esse isolamento fosse prolongado no tempo e tivesse o sucesso funcional desejado. Tal circunstância não implicaria qualquer divergência evolutiva, uma vez que a compatibilidade reprodutiva não seria alterada. Introduzamos agora um novo elemento neste hipotético cenário de desenvolvimento: uma alteração genética é produzida no seio de um conjunto dessas populações expostas previamente a um isolamento geográfico e/ou reprodutivo. As pressões seletivas favoreceriam essa mutação, que finalmente poderia vingar. Verifica-se aqui um processo saltacionista? Só se a mutação tivesse lugar em diferentes indivíduos ou fosse facilmente transmissível, originando um efeito “fundador” dessa divergência (neste caso, através da dominância ou recessividade genética inerente à mutação em causa). A probabilidade deste fenómeno acontecer é potencialmente muito pequena, contudo são também diversas as variáveis que interferem nessa possibilidade mutagénea17, 18, bem como o tipo de mutação realizada19. Porém, o sucesso dessas populações concentrar-se-á também no seu desenvolvimento morfogenético, pois só dessa forma seria viável o resultado dessa mutação (qualquer mudança só tem sentido se é funcionalmente aproveitável pelo sistema). Sendo assim, o saltacionismo associado ao pool genético sustentar- se-ia na sua capacidade de reproduzir variabilidade fenotípica, genética e ambiental (esta última sujeita à própria diversificação funcional que a variabilidade de novas respostas proporcionaria). Essa capacidade de desenvolvimento geraria o gradualismo de “esperanças monstruosas”, neste caso concentradas em fenótipos descontínuos isolados reprodutivamente20, ou não (sempre de acordo com a facilidade de manter o fluxo reprodutivo original).
Ambos os exemplos são úteis para associar dois mecanismos potencialmente divergentes, que acabem por canalizar processos de alteração dentro de cada plano corporal. Porém, surge aqui uma outra interrogante: acaso esse processo conjunto gradualista/ saltacionista estaria sujeito a uma linearidade própria de cada plano corporal? Isto é, a diferença entre processos graduais e pontuais (saltacionistas) sustentar-se-ia sobre hipotéticas linearidades ou não-linearidades no desenvolvimento e evolução? Dito doutra forma, e assumindo a existência de um hipotético plano corporal, seria factível detetarmos uma tendência comportamental nesse processo?
A escala analítica — linearidades e não-linearidades
A linearidade/não-linearidade descreve-se com base na perspetiva aplicada. Como já vimos, a perspetiva macroanalítica define supostos planos corporais e, por esse motivo, aparentes linearidades, no que a desenvolvimento e evolução diz respeito. Não devemos esquecer que o plano corporal constitui o conjunto de características morfogenéticas partilhadas, conservadas e potenciadas pelos membros de um filo21. Um dos principais argumentos para esta ideia é a assimetria funcional dos organismos, i.e., alguns órgãos ou partes parecem ter maior relevância do que outros, menos suscetíveis de evoluir e contribuir para planos corporais específicos22. Esses carateres mais relevantes constituiriam key innovations23, atendendo à capacidade de criar conjuntos de respostas fenotípicas (desenvolvimento) que contribuíssem para o funcionamento dos sistemas. Conforme descemos na perspetiva aplicada para a análise morfogenética dos indivíduos, essa linearidade vai perdendo consolidação (como registamos na FIGURA 1), aumentando a desordem morfogenética e, portanto, a hipotética consolidação de planos corporais. É esse aumento considerável da desordem que faz com que a linearidade ceda espaço à não-linearidade.
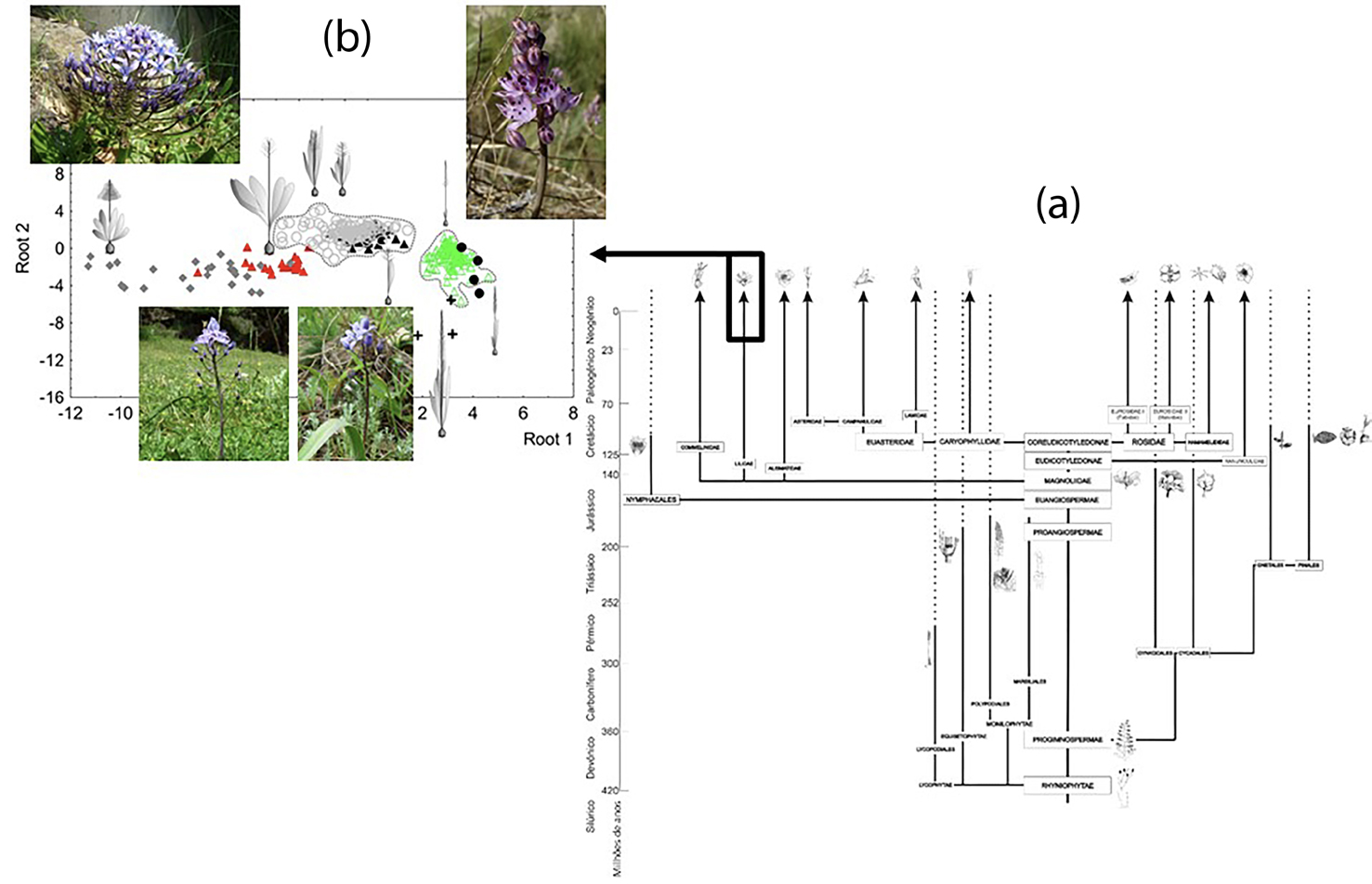
Os planos corporais constituem, por este motivo, um instrumento de utilidade crescente conforme avançamos na escala analítica (de micro a macro). Neste âmbito, a linearidade ganha protagonismo e, assim, é possível descrever linhas filogenéticas. Por este motivo, saltacionismo e gradualismo estarão igualmente sujeitos a essa escala analítica, ganhando consolidação ou perdendo-a, conforme a perspetiva envolvida (mais macro ou mais microanalítica, respetivamente).
Referências
- 1 KUTSCHERA, U. & NIKLAS, K. J., Darwin-Wallace Demons: survival of the fastest in populations of duckweeds and the evolutionary history of an enigmatic group of angiosperms, Plant Biology, Plant Biology, 17, p 24-32. 2015.
- 2 NIKLAS, K. J. & KUTSCHERA, U., The evolutionary development of plant body plans, Functional Plant Biology, Functional Plant Biology, 36, 8, p 682-695. 2009.
- 3 THEISSEN, G., Saltational evolution: hopeful monsters are here to stay, Theory in Biosciences, Theory in Biosciences, 128, 1, p 43-51. 2009.
- 4 RACOVSKI, T., Evolutionary novelty: a philosophical and historical investigation, Thesis for the degree of Doctor of Philosophy in Philosophy, University of Exeter. 2019.
- 5 NOVICK, A., Cuvierian functionalism, Philosophy, Theory, and Practice in Biology, 11. 2019.
- 6 HEARD, S. B. & HAUSER, D. L., Key evolutionary innovations and their ecological mechanisms, Historical Biology, 10, 2, p 151-173. 1995.
- 7 HUNTER, J. P., Key innovations and the ecology of macroevolution, Trends in Ecology & Evolution, 13, 1, p 31-36. 1998.
- 8 WAGNER, G. P., The origin of morphological characters and the biological basis of homology, Evolution, 43, 6, p 1157- 1171, 1989.
- 9 LOVE, A. C., Functional homology and homology of function: Biological concepts and philosophical consequences, Biology & Philosophy, 22, 5, p 691-708. 2007.
- 10 HALLGRÍMSSON, B. et al., The generation of variation and the developmental basis for evolutionary novelty, Journal of Experimental Zoology Part B: Molecular and Developmental Evolution, 318, 6, p 501-517. 2012.
- 11 SCHWARTZ, J. H., Darwinism versus Evo–Devo: a late–nineteenth century debate, A Cultural History of Heredity III: 19th and Early 20th Centuries, 67. 2005.
- 12 PETROV, R. V. & VOROBYEVA, E. I., Fundamentals of biological science: an evolutionary approach, Physiological and Health Sciences in the global Encyclopedia of Life Support Systems (EOLSS). 2009.
- 13 PIGLIUCCI, M. & MÜLLER, G. B., Elements of an extended evolutionary synthesis, MIT Press, p 3-17. 2010.
- 14 PIGLIUCCI, M., Do we need an extended evolutionary synthesis?, Evolution: International Journal of Organic Evolution, 61, 12, p 2743-2749. 2007.
- 15 ANTONELLI, C., The system dynamics of collective knowledge: From gradualism and saltationism to punctuated change, Journal of Economic Behavior & Organization, 62, 2, p 215-236. 2007.
- 16 WEST-EBERHARD, M. J., Developmental Plasticity and Evolution, Oxford University Press USA, New York, 793 pp. 2003.
- 17 OTTO, S. P. & WHITLOCK, M. C., The probability of fixation in populations of changing size, Genetics, 146, 2, p 723-733, 1997.
- 18 WAHL, L. M. & GERRISH, P. J., The probability that beneficial mutations are lost in populations with periodic bottlenecks, Evolution, 55, 12, p 2606-2610. 2001.
- 19 BOUZAT, J. L. et al., Beyond the beneficial effects of translocations as an effective tool for the genetic restoration of isolated populations, Conservation Genetics, 10, 1, p 191-201. 2009.
- 20 LEVIT, G. S. & HOSSFELD, U., Darwin without borders? Looking at ‘generalised Darwinism’ through the prism of the ‘hourglass model’, Theory in Biosciences, 130, 4, p 299-312. 2011.
- 21 IRIEL, N. & KURATANI, S., The developmental hourglass model: a predictor of the basic body plan?, Development, 141, p 4649-4655. 2014.. DOI: 10.1242/dev.107318.
- 22 WILLMORE, K. E., The body plan concept and its centrality in Evo-Devo, Evolution: Education and Outreach, 5, p 219- 239. 2012.
- 23 MÜLLER, G. B. & PAGEL, M. D., Novelty and key innovations, Encyclopedia of evolution, 2, p 827-830. 2002.
- 24 ALMEIDA DA SILVA, R. et al., The Iberian species of Scilla (subfamily Scilloideae, family Asparagaceae) under climatic change scenarios in southwestern Europe, Systematic Botany, 39, 4, p 1083-1098. 2014.
Este artigo já foi visualizado 3277 vezes.