

Breve guia de Paleontologia urbana
Fósseis de cefalópodes
📧 , 📧
- * DG/ FC/ U. Lisboa
- ɫ CG/ DCT/ U. Coimbra
Referência Silva, C. M., Pereira, S., (2026) Breve guia de Paleontologia urbana, Rev. Ciência Elem., V14(1):008
DOI http://doi.org/10.24927/rce2026.008
Palavras-chave
Os cefalópodes, da imaginação à realidade.
São vários os grupos biológicos que, estimulando a sugestionável imaginação humana, geraram figuras mitológicas, fantásticas. Contudo, quimeras, grifos, unicórnios, harpias, dragões, hidras, o Pégaso – entre muitas outras – são todas, de algum modo fabuloso e desconcertante, organismos vertebrados. Entre os invertebrados apenas os cefalópodes – e alguns artrópodes como o infame crustáceo Carcino, ou Câncer – habitando as insondáveis profundezas marinhas, têm potencial para gerar, mas só na nossa mente, seres simultaneamente prodigiosos e temíveis.
O Kraken, criatura tremenda capaz de gelar o sangue do mais audaz marinheiro e de arrastar, sem dó, nem piedade, navios inteiros para as assombrosas profundezas do Mar do Norte, era descrito como um polvo gigantesco e aterrador. Conseil, o criado do Professor Aronnax, a bordo do Nautilus nas 20.000 Léguas Submarinas de Jules Verne, fala dele com apreensão. O mítico submarino do Capitão Nemo, o Nautilus, o navegador, foi nomeado em honra de outro cefalópode, mas este bem real.
Olhando apenas para os cefalópodes da atualidade, para as lulas e os polvos deste final de Cenozoico, seria difícil imaginar quão mais diversificado este importante grupo de moluscos foi no passado. Seria uma tarefa igualmente desafiante entender a sua vasta representação no registo fóssil do Fanerozoico, em rochas com idades que variam entre os cerca de 538 e os 66 milhões de anos (Ma) de idade, do Câmbrico ao final do Cretácico, e qual a razão da sua extrema importância para a biostratigrafia, ou seja, para a datação das sequências de estratos de génese marinha do Paleozoico e do Mesozoico.
Esta circunstância deve-se ao facto de a esmagadora maioria dos cefalópodes atuais não possuírem partes esqueléticas mineralizadas, conchas duras. Mesmo os chocos, outro grupo de cefalópodes que nos é bem familiar, apresentam esqueleto interno delicado, algo frágil – o sepion, ou osso de siba – e, por isso mesmo, menos propício à fossilização. Consequentemente, os fósseis de chocos são escassos e os de polvos extremamente incomuns.
Contudo, na atualidade existe um outro grupo de cefalópodes, os náutilos, que nos poderão ajudar a esclarecer o aparente paradoxo entre as características pouco favoráveis à fossilização da maioria dos cefalópodes atuais e a sua ampla representação no registo fóssil. Os náutilos, hoje em dia representados pelos géneros Nautilus e Allonautilus, totalizando umas nove espécies, cerca de 10% da diversidade dos cefalópodes atuais, ostentam concha externa biomineralizada, resistente. Além disso, existiram no passado outros grupos de cefalópodes, elementos cruciais dos ecossistemas marinhos de então que, entretanto, se extinguiram, desaparecendo para sempre da face do planeta. É o caso dos amonoides, cujos representantes mais conhecidos são as amonites, para além de muitos outros.
Daí que não seja incomum encontrar vestígios fossilizados destes seres preservados nos elementos da geodiversidade utilizados1 nas construções das nossas cidades, nas rochas ornamentais das fachadas e pavimentos dos edifícios. No caso das amonites, cefalópodes emblemáticos do Mesozoico, até os podemos ver como motivos artísticos, embelezando as paredes exteriores de museus (FIGURA 1 A)). Em Portugal, são menos comuns que os fósseis dos rudistas2, é um facto, mas estão lá! Basta – saber – procurá-los! Estão também nos pavimentos e nas paredes urbanas um pouco por todo o mundo (FIGURA 1 B)). Assim, o propósito deste segundo opúsculo do Breve Guia de Paleontologia Urbana é, precisamente, ajudar a reconhecê- los e a interpretá-los.

Origem e evolução dos cefalópodes.
Para entender que fósseis de moluscos cefalópodes encontramos em meio urbano e porquê, temos de entender, por exemplo, como eles se originaram e diversificaram, desde o Câmbrico até à atualidade (FIGURA 2). Também teremos de compreender por que ocorrem nas rochas em que os encontramos, rochas essas que são um registo dos ambientes em que viviam. Assim sendo, o que são os cefalópodes e como se originaram? E onde vivem?
Os cefalópodes são organismos marinhos, ocorrendo a diversas profundidades, desde a superfície do oceano até às profundezas abissais. Polvos e chocos podem ainda ocorrer em ambientes estuarinos. São pescados no estuário do Tejo, por exemplo3. Parte significativa deles – lulas e náutilos – tem modo de vida pelágico nectónico, nadando na coluna de água dos oceanos. Os polvos, contudo, são organismos demersais, isto é, mantêm a capacidade de nadar, mas vivem na dependência do fundo, abrigando-se nele e caçando presas bentónicas que lá vivem. O mesmo, em termos gerais, para os chocos. Todos são carnívoros, predadores ativos, possuindo braços – ou tentáculos, ou ambos – munidos de ganchos ou de ventosas, dotados de bicos córneos na boca e providos de olhos bem desenvolvidos. Os cefalópodes deslocam-se por meio de um sistema de propulsão a jato de água, expelindo água aspirada para o interior da cavidade do manto através do hipónoma, do funil do corpo mole (FIGURA 3). Têm em comum com os restantes moluscos, com os bivalves e os gastrópodes, por exemplo, o facto de serem organismos com simetria bilateral e possuírem um órgão, o manto, responsável, nomeadamente, por segregar a concha calcária, quando presente. Entre os cefalópodes contam-se os maiores invertebrados conhecidos, a lula gigante Architeuthis dux, podendo atingir 13 m de comprimento total4, e aqueles que – tento um sistema nervoso central sofisticado, para um animal invertebrado – são tidos como mais inteligentes, ou seja, que manifestam comportamentos mais complexos e demonstram capacidade de aprendizagem mais elevada. Sim, é possível ensinar “truques” a um choco5!
No modelo lineano de organização da biodiversidade atualmente aceite, os Cephalopoda constituem um grupo biológico, um táxone, de categoria classe no seio do Filo Mollusca, o grupo dos moluscos. O termo, do grego ϰεφαλή kephalé + ποδός podós, significando cabeça e pé, foi cunhado em 1795 pelo grande naturalista francês Jean Léopold Nicolas Frédéric Cuvier, conhecido como Georges Cuvier (1769–1832), o famoso “Pai da Paleontologia”. Cuvier iniciou a sua caracterização do grupo, curiosamente, com: “Leur téte est libre, mobile et couronnée de grands tentacles sur lesquels ils marchent. (…)”6, seguramente aludindo ao polvo.
Entre os cefalópodes atuais reconhecem-se dois subgrupos, duas subclasses: Coleoidea, incluindo os polvos, as lulas e os chocos, e Nautiloidea, os náutilos. Estão representados nos ecossistemas marinhos da atualidade por cerca de 800 espécies. No passado, os cefalópodes incluíam outras subclasses, hoje extintas. A mais conhecida é a dos Ammonoidea, os amonoides, que existiram do Devónico Inferior, desde os 410 Ma, até aos finais do Cretácico, até aos 66 Ma.
A história destes moluscos é extremamente antiga. É possível que remonte aos primórdios do Fanerozoico, aos inícios do Câmbrico (FIGURA 2). Contudo, os mais antigos registos fidedignos destes organismos são umas dezenas de milhões de anos mais recentes, datando dos finais do Câmbrico7. Estes primeiros cefalópodes eram animais de pequenas dimensões, entendidos como sendo representantes dos Nautiloidea s.l. Possuíam conchas externas cónicas rectilíneas ou levemente encurvadas, subdivididas internamente em câmaras, medindo poucos centímetros de comprimento. Estes organismos nadadores, presume-se, seriam descendentes de outros moluscos, mas bentónicos, vivendo nos fundos marinhos. Provavelmente, de monoplacóforos em que a concha cónica foi modificada, transformando-se num dispositivo de flutuação provido de câmaras internas individualizadas por tabiques ou septos8. Foi esta concha externa tabicada, funcionando como aparelho de flutuação, que permitiu aos primeiros cefalópodes libertar-se dos habitates bentónicos dos fundos marinhos e povoar ambientes da coluna de água como organismos pelágicos nadadores.
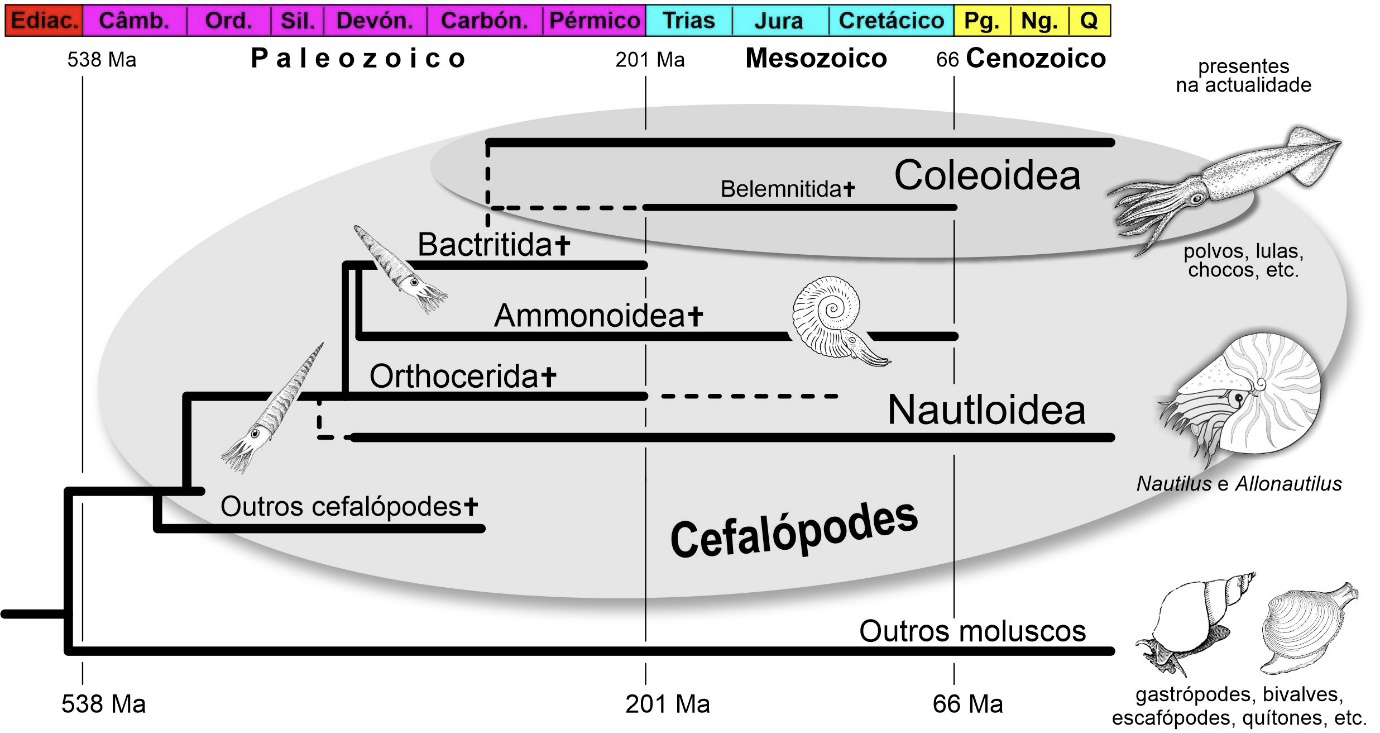
Os nautiloides s.l. – incluindo, entre outros, os Endoceratoidea, os Actinoceratoidea e os Nautiloidea, ainda que não exista consenso quanto a esta repartição – experimentaram uma ampla diversificação a partir do início do Ordovícico, atingindo o seu máximo de diversidade durante o Paleozoico. É também nesta altura que atingem maiores dimensões. A concha de alguns exemplares do nautiloide ortocónico ordovícico Cameroceras alcançava frequentemente 6 m de comprimento!9, podendo alguns exemplares chegar aos 10 m!10 De seguida, sofrem redução significativa como resultado do evento biótico global do final do Pérmico, aos 250 Ma. Apenas os Nautiloidea s.s. sobreviveram a esta extinção em massa, existindo no Mesozoico e chegando, à atualidade, ainda que vestigialmente.
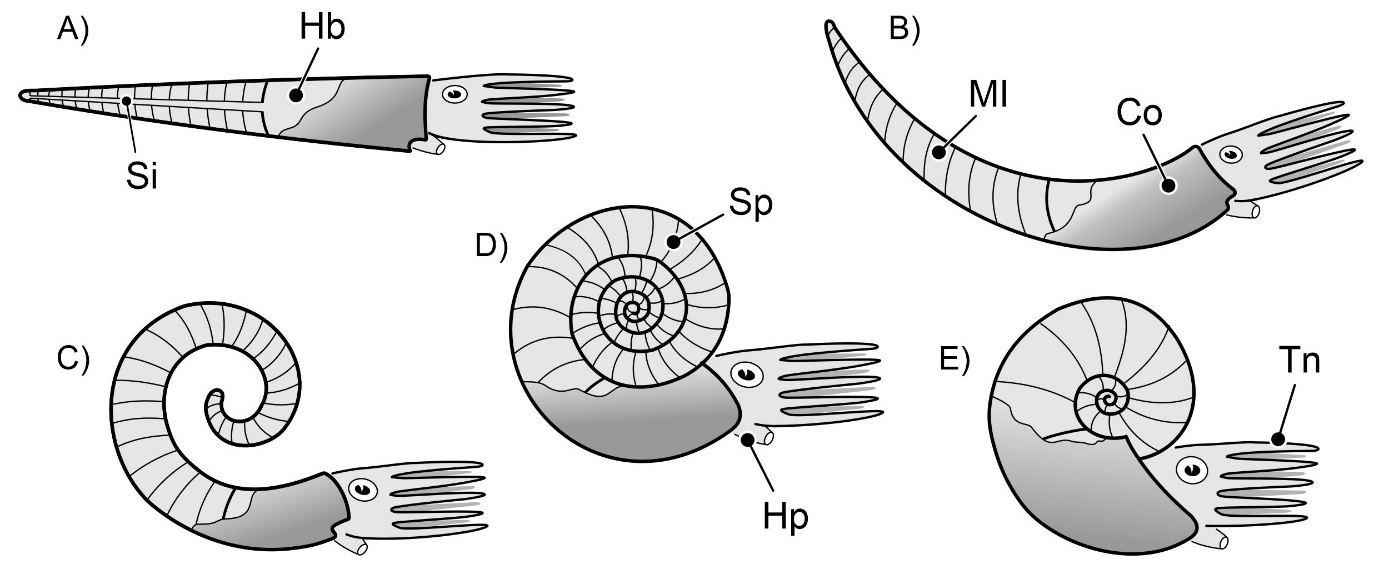
Se do Mesozoico à atualidade os nautiloides que predominaram tinham conchas externas tabicadas fortemente espiraladas, nautilicónicas, como em Nautilus, durante o Paleozoico a forma das conchas era bastante mais variada, raramente espiralada com espira apertada (FIGURA 3E). As mais comuns eram as ortocónicas, em forma de cone retilíneo (FIGURA 3A), como em Orthoceras, ou levemente arqueadas, cirtocónicas (FIGURA 3B).
Nos mares da atualidade, os coleoides, Classe Coleoidea, são os cefalópodes mais abundantes, perfazendo 99% da diversidade total do grupo. Encontram-se amplamente distribuídos por todos os oceanos, dos polos aos trópicos e de ambientes litorais a abissais. Polvos e chocos ocorrem também em estuários. Anatomicamente, distinguem-se por possuírem uma concha interna reduzida (nos chocos) ou vestigial (lulas), podendo nem apresentar esqueleto biomineralizado (polvos). Possuem 10 a oito apêndices – braços e ou tentáculos – associados à boca, circum-orais, i.e., circundando-a.
A origem dos coleoides remonta ao Paleozoico. Terão tido como antepassado um grupo de cefalópodes distinto, os Bactritida8, dotados de concha externa ortocónica a cirtocónica (FIGURA 2). Fósseis do Devónico Inferior foram interpretados como representando coleoides, contudo, vários autores contestam a classificação destes exemplares. Os espécimes fossilizados mais antigos atribuídos incontestavelmente a este grupo datam do Paleozoico superior, do Carbónico11, tendo cerca de 325 Ma. Tratando-se de animais com elementos biomineralizados reduzidos ou ausentes, o seu registo fóssil é – com a exceção das belemnites – escasso, o que dificulta bastante o estudo da sua origem e evolução.
Os polvos, um outro grupo de cefalópodes coleoides bem representado na atualidade, terão surgido no Jurássico Inferior8. Estes animais não possuem esqueleto biomineralizado, concha dura, consequentemente, os fósseis de polvos são extremamente incomuns. O mais famoso exemplar fossilizado de um destes animais, devido à preservação excecional dos seus tecidos moles, é seguramente o de Proteroctopus ribeti, do Jurássico Médio de França12.
Os coleoides mais bem representados no registo fóssil são as belemnites, pertencendo à ordem Belemnitida, um grupo de animais com aspeto similar ao das atuais lulas. Contudo, ao contrário das lulas, as belemnites são um grupo extinto, inteiramente mesozoico. Além disso, possuíam esqueleto interno biomineralizado, com um elemento bastante resistente, o rostro, daí a sua abundante representação no registo paleontológico.
Um grupo de cefalópodes extremamente importante são os amonoides. Contudo, tratando-se de um táxon extinto há 66 milhões de anos, não nos é familiar como o são as lulas, os polvos ou até mesmo os náutilos. Os amonoides constituem a Subclasse Ammonoidea dos cefalópodes. Eram organismos dotados de concha externa tabicada, ou seja, dividida internamente em câmaras por meio de septos, de tabiques, tal como as dos náutilos. O facto de possuírem concha resistente torna a probabilidade de fossilizarem muito mais elevada. Daí os seus fósseis serem comuns, o que é excelente para o seu estudo. É complicado estudar coisas raras.
Os amonoides surgiram no Paleozoico, no início do Devónico, há cerca de 410 Ma, tendo como antepassados os Bactritida que, por sua vez, evoluíram de cefalópodes nautiloides Orthocerida (=Michelinocerida)8 surgidos no Ordovício Inferior (FIGURA 2). Extinguiram-se no final do Mesozoico. Foram vitimados, ao que tudo indica, pelo mesmo evento biótico de extinção em massa do final do Cretácico que varreu da face da Terra os dinossáurios não-avianos em ambientes continentais e os rudistas2 em ambientes marinhos, entre muitos outros grupos biológicos extraordinários, quer de animais, quer de plantas13, 14, 15.
Ao longo da sua existência, os amonoides diversificaram-se imenso, dando origem a muitos subgrupos de categoria ordem. Os três mais conhecidos são os Goniatitida, que surgiram no Devónico Médio há uns 390 Ma e se extinguiram no final do Pérmico, há cerca de 250 Ma, os Ceratitida, que existiram do Pérmico ao final do Triásico, e os Ammonitida, do Jurássico ao Cretácico (FIGURA 2). Talvez a característica distintiva mais evidente destes grupos seja as suas linhas de sutura, isto é, o contorno do bordo dos septos internos na união com a face interior da parede externa da concha, visível à superfície dos seus moldes internos. Distinguem-se linhas goniatíticas, ceratíticas e amoníticas, entre outras. É importante notar que, sendo – por exemplo – a linha de tipo ceratítico mais comum entre os amonoides Ceratitida, ela também pode ocorrer em amonoides mais recentes, em amonites. O mesmo para outros tipos de linhas. A natureza é complexa! Só é simples na mente dos humanos.
Ainda que os cefalópodes sejam um elemento muito importante dos ecossistemas marinhos da atualidade, quer pela sua abundância, que pela diversidade, é importante reter que, no passado, como vimos, incluíam muitos mais grupos que hoje (FIGURA 2). A diversidade de cefalópodes que conhecemos atualmente, ainda que grande, é apenas uma pequena amostra da variedade de espécies destes organismos que existiram no passado, desde os inícios do Paleozoico até hoje.
A biodiversidade do planeta, como um todo, não inclui apenas a que existe hoje. Engloba também a biodiversidade do passado, deste o início da vida sobre a Terra, há mais de 3500 Ma. Sabemos que essa paleobiodiversidade existiu, tendo dado origem à diversidade biológica que conhecemos na atualidade, porque encontramos hoje os vestígios fossilizados dos representantes desses grupos biológicos nos estratos rochosos da crosta terrestre. Esses fósseis são elementos da geodiversidade1, da componente abiótica da natureza. Só investigando a história da vida do passado, estudando os seus vestígios fossilizados, entenderemos a origem da biodiversidade do presente e estaremos em condições de promover a salvaguarda da vida do futuro.
Fossilização. Fenómeno raro?
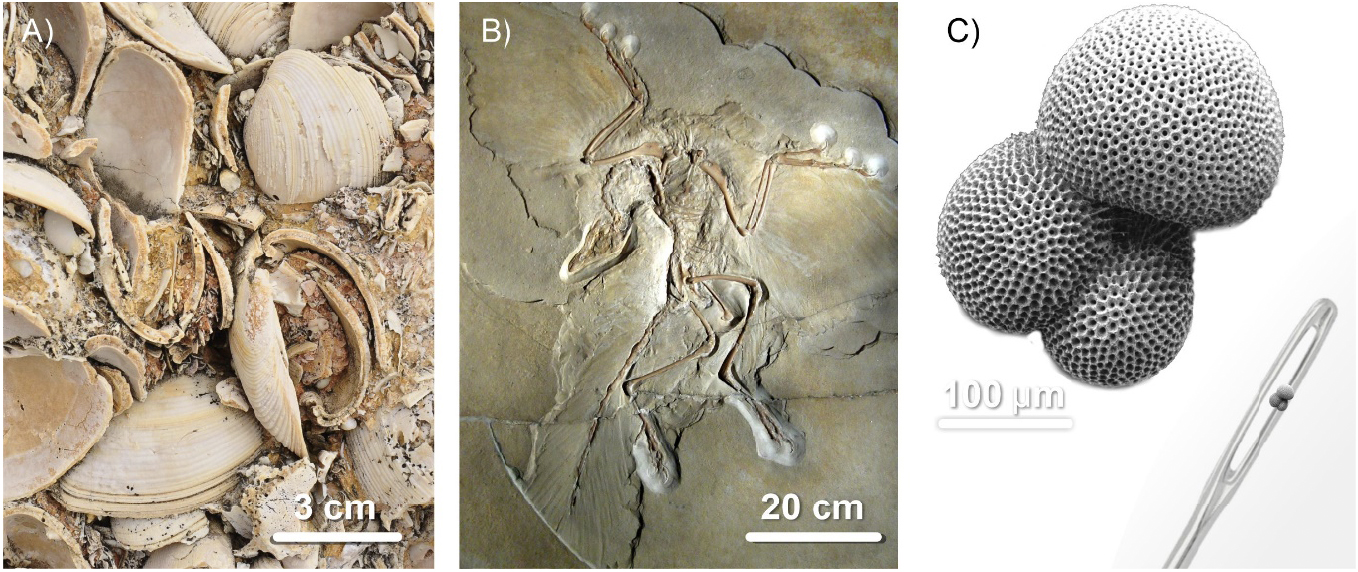
Ao contrário do que frequentemente se pensa, a fossilização, a formação de fósseis, é um fenómeno extremamente comum. Há fósseis um pouco por todo o lado, sobretudo em estratos de rochas sedimentares, mas não só. Há até rochas que são formadas, na sua maioria, por restos fossilizados de organismos do passado. São as rochas sedimentares biogénicas, constituídas por mais de 50% de bioclastos, de fragmentos de conchas e de carapaças (FIGURA 4). Por vezes, os fósseis são tão pequenos – microfósseis – que nem os vemos. Microscópicos. Mas estão lá!
Para gerar fósseis não são necessárias condições extraordinárias. As condições ditas “especiais”, como privação de oxigénio, ausência de transporte pós-morte ou enterramento rápido, por exemplo, são necessárias, isso sim, para obter fósseis excecionais. Por exemplo, para se conseguir fossilizar elementos do corpo mole dos organismos, elementos de natureza orgânica, é benéfico que esses restos sejam preservados em ambientes desprovidos de oxigénio, anóxicos. Para se obter fósseis de esqueletos articulados, é vantajoso que o cadáver do animal não seja arrastado e desmembrado após a morte, sendo soterrado rapidamente15. De facto, assim é! Mas para obter um fóssil da concha resistente de um bivalve ou de um gastrópode marinho, de um búzio, nada disto é necessário. As condições que observamos normalmente na praia, onde encontramos acumulações de conchas na zona exposta na maré-baixa, são o suficiente. Por isso há milhões de fósseis de conchas de bivalves, de trilobites e de amonites e tão poucos de polvos, de medusas e de vermes.
E depois ainda há a categoria dos icnofósseis, dos fósseis de vestígios de atividade orgânica. Senão vejamos, os somatofósseis de vermes, i.e., os fósseis de elementos do seu corpo, uma vez que não possuem partes biomineralizadas, são muito invulgares. Mas os seus icnofósseis, os fósseis dos túneis escavados por eles no sedimento dos fundos marinhos, por exemplo, são extremamente comuns!
Fala-se hoje muito de “perceções”. O que leva à perceção de que a fossilização é um fenómeno raro e que os fósseis são escassos e, como tal, difíceis de encontrar é a fixação do público nos vestígios dos animais vertebrados. Mais, é o fascínio pelos fósseis de vertebrados apresentando o esqueleto articulado, referido muitas vezes como “completo”. Esses sim, são incomuns. Contudo, não se deve confundir a raridade desses fósseis conservando aspetos extraordinários com a raridade da fossilização, em geral. Ainda que sejam fenómenos relacionados, são circunstâncias distintas.
Outro aspeto que altera a perceção que se tem da fossilização é a chamada infidelidade do registo fóssil. A “incompleteness of the fossil record” dos autores anglo-saxónicos. Este fenómeno resulta do facto de a fase inicial da fossilização, a fase biostratonómica, aquela que ocorre entre a morte dos organismos – ou a geração dos restos passíveis de fossilizar – e o seu enterramento, ser muito exigente do ponto de vista mecânico, do ponto de vista da resistência física dos vestígios a fossilizar. É uma etapa marcada pela degradação dos tecidos moles, pela desarticulação dos esqueletos e pelo desgaste e fragmentação dos restos daí resultantes. Tudo isto ocorre como resultado de transporte, isto é, do arrastamento pós-morte a que estão muito frequentemente sujeitos os cadáveres e os restos orgânicos, seja por agentes biológicos ou físicos.
Os restos de organismos sem partes esqueléticas resistentes, duras, têm uma capacidade mais baixa de resistir à fase biostratonómica e, como tal, de serem incorporados no registo fóssil, de fossilizar. Do mesmo modo, os fósseis articulados de vestígios biológicos constituídos por diversos elementos esqueléticos independentes, por valvas, ossos, artículos, espículas, etc., são significativamente mais incomuns que os fósseis isolados desses mesmos elementos.
Contudo, mais uma vez, o facto de muitos grupos de organismos, os de corpo mole, orgânico, estarem sub-representados no registo geológico não significa que a fossilização seja um fenómeno raro. Incorporados na crosta terrestre há milhões e milhões de fósseis de conchas, de carapaças, de troncos, de dentes, de espinhos e até de pegadas e outros vestígios de muitos organismos diferentes, quer de animais, quer de plantas. A fossilização não é rara, é seletiva.
Há tantos fósseis, sobretudo nas rochas sedimentares, que até os encontramos nas fachadas e nos pavimentos das nossas cidades. Nomeadamente, fósseis de cefalópodes. Eles estão lá. Umas vezes maiores, outras mais pequenos. Umas, mais bem preservados, outras, mais fragmentados. Nós é que – apressados a caminho do trabalho ou da escola – não lhes damos a devida atenção.
Que fósseis de cefalópodes encontramos na cidade e como os identificar?
Os fósseis, juntamente com as rochas, os minerais, os vulcões, os recursos minerais, os solos, etc., etc., são elementos da geodiversidade. São constituintes da parte abiótica, não viva, da natureza. Os materiais geológicos, sobretudo as rochas, são trazidas para a cidade, para a construir. As nossas casas, as ruas, os monumentos, os edifícios públicos, os castelos e os palácios, etc., são construídos com materiais geológicos, com pedra de construção e com rochas ornamentais. É nesses materiais que encontramos os fósseis em meio urbano, nomeadamente, os de cefalópodes (FIGURA 5).

Em Portugal, e por esse mundo fora, em contexto citadino, mais frequentemente podemos encontrar vestígios de três grupos de cefalópodes: de coleoides e de nautiloides e, ainda, de amonoides, extintos desde o final do Mesozoico (FIGURA 5).
No nosso país, em contexto de geodiversidade urbana – que, tal como o salientamos quando se abordaram os fósseis de rudistas, não deve ser confundido com ocorrências naturais, nem a elas igualado, – a maioria dos fósseis de cefalópodes surge associada a rochas ornamentais de idade mesozoica, Jurássico e Cretácico, pois são aquelas que pelas suas características físico-mecânicas e estéticas melhor se adequam à construção. Por exemplo, em calcários Azul Valverde, do Jurássico Superior18 da região de Alcanede, Santarém, Vidraço de Moleanos e Ataíja, Azul e Creme, também jurássicos, ainda que mais antigos, Jurássico Médio, da região de Alcobaça, Leiria, do Maciço Calcário Estremenho. Mas não só!
Curiosamente, nas nossas cidades, pelo menos em Lisboa (no pavimento do Terreiro do Paço e nos de outros locais), Setúbal (Convento de Jesus, Sala do Capítulo) e Alcácer do Sal (Igreja do Espírito Santo, Museu Municipal Pedro Nunes), por exemplo, também ocorrem fósseis de cefalópodes associados a rochas exóticas, importadas de longe. É o caso do calcário fossilífero Folkeslunda de Öland, na Suécia1, também conhecido como “calcário de Orthoceras”. Esta rocha é bem mais antiga que os calcários nacionais referidos. É de idade Ordovícico, Paleozoico19.
No calcário cretácico Liós da região de Lisboa, rico de fósseis de rudistas, não são conhecidos vestígios de cefalópodes. Os ambientes de plataforma pouco profunda, junto à costa, onde proliferavam bancos de rudistas, não eram propícios à existência de animais pelágicos nectónicos, de animais que nadassem na coluna de água, já que – por exemplo – essa coluna de água era pouco significativa. Não será impossível encontrar fósseis de cefalópodes no Liós. Contudo, essa ocorrência é bastante improvável.
Como visto anteriormente, no caso dos fósseis de rudistas, alguns dos elementos de identificação dos seus fósseis estão patentes nas secções expostas em paredes e pavimentos urbanos1. Para os cefalópodes, contudo, os elementos de identificação dos diversos grupos, mesmo dos de categoria mais elevada, categoria Ordem, por exemplo, tais como Ceratitida, Ammonitida, etc., estão — na sua maioria — apenas patentes à superfície tridimensional dos moldes internos ou das mineralizações das conchas. Esses elementos diagnósticos não são visíveis em secções. Isto torna difícil — mas não completamente impossível — a identificação dos seus fósseis em fachadas e pisos. Tal como referido anteriormente1, é fundamental ter-se em conta que a identificação de fósseis em secção expostos em contexto urbano, não é uma ciência exata. Trata-se de uma abordagem expedita tendo em vista a divulgação científica e o usufruto dos fósseis que ocorrem na cidade, não de uma análise paleontológica convencional em condições laboratoriais ótimas. É a abordagem — e a identificação — possível, não a ideal.
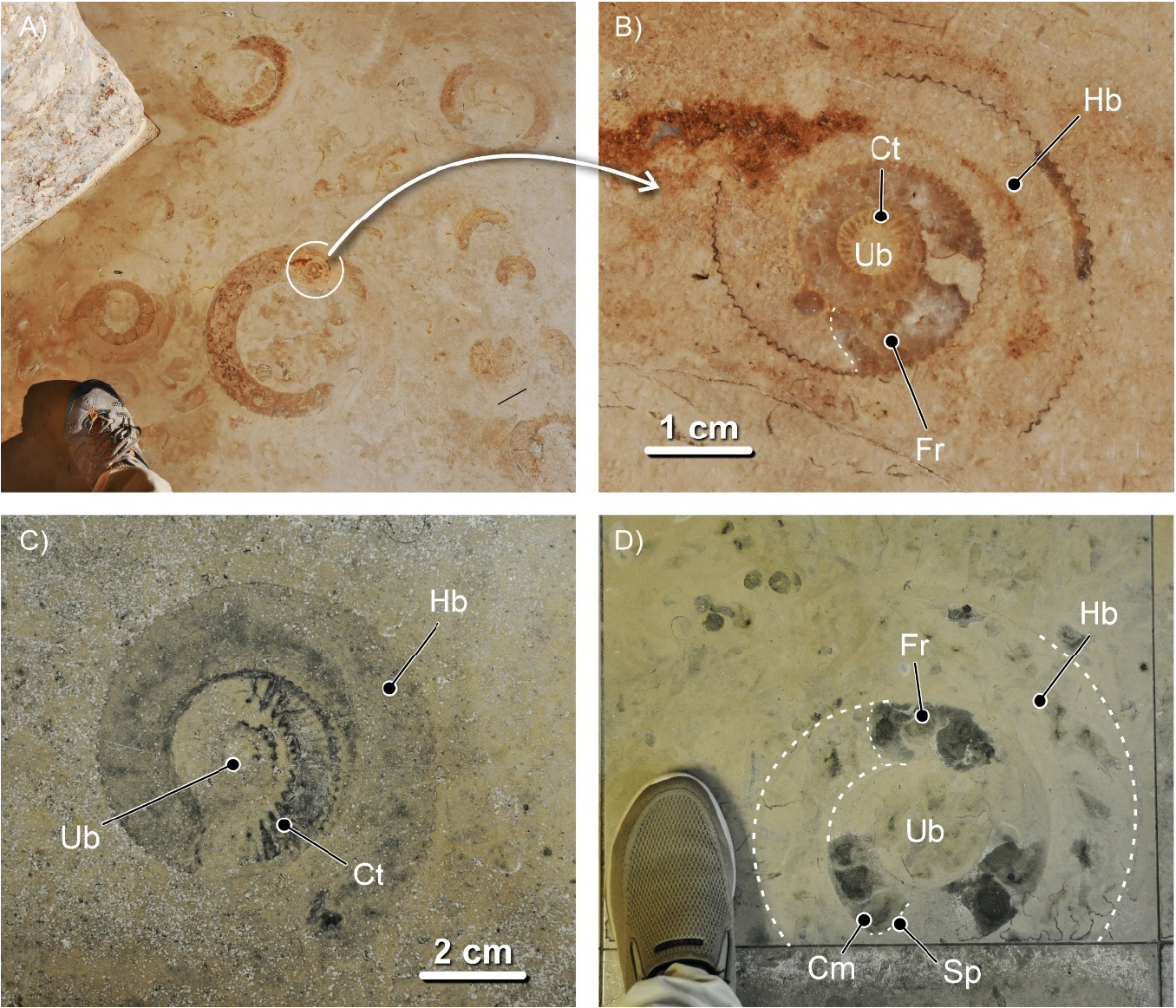
Uma vez que os fósseis de cefalópodes em contexto urbano surgem em secções bidimensionais expostas na superfície de fachadas e em pavimentos, para os identificar e interpretar adequadamente há que ter em conta a orientação dos cortes que as originaram, quer para os nautiloides (e.g., FIGURA 6), quer para os amonoides e as belemnites.
Os nautiloides e os seus fósseis na cidade.
Não obstante os amonoides estarem extintos há muito, nautiloides e amonoides apresentam vários aspetos em comum, uma vez que são grupos de organismos aparentados, sendo os amonoides primos afastados dos nautiloides (FIGURA 2). Contudo, também apresentam diferenças significativas que justificaram a sua separação em grupos independentes.
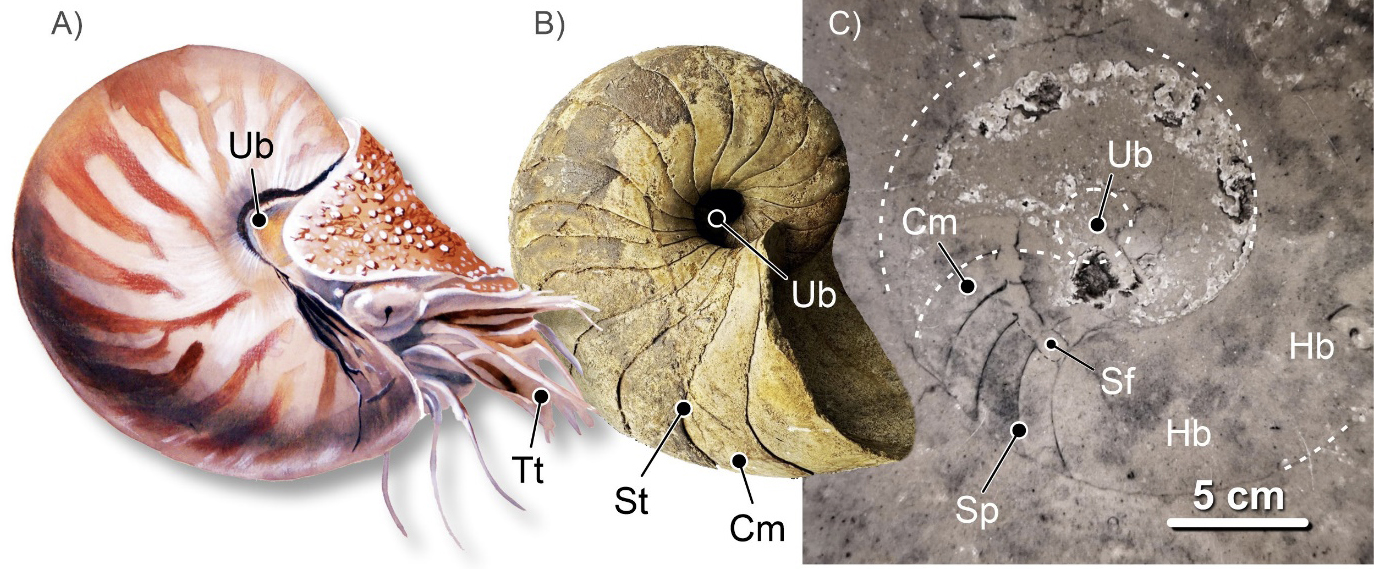
Em ambos os grupos, os seus representantes desenvolveram conchas biomineralizadas externas, subdivididas internamente em câmaras, por meio de septos ou tabiques. Por isso se diz que o Nautilus da atualidade possui concha externa tabicada. A porção da concha que apresenta septos, da câmara inicial – no início do crescimento da concha do animal – até ao último septo, é denominada fragmocone (FIGURA 7). A última câmara, ocupada pela maior parte do corpo visceral do animal, é denominada câmara de habitação ou habitáculo. O animal controla o que se passa em todos os espaços interiores da concha. Nomeadamente, nas câmaras do fragmocone, por via de um elemento do seu corpo mole, o sifúnculo. É assim que o Nautilus regula a flutuabilidade do seu corpo na coluna de água, consoante – ao nadar – mergulha ou emerge. O sifúnculo estava alojado numa estrutura carbonatada da concha, por vezes muito complexa, no sifão. O sifúnculo, sendo de natureza orgânica, não se preserva nos fósseis. Mas o sifão, esse, está frequentemente bem patente (FIGURA 6C e 7C).
Ao padrão linear resultante da união da periferia dos septos do fragmocone com a parede externa da concha, padrão esse só visível nos fósseis à superfície dos moldes internos da concha, dá-se o nome de linha de sutura. Este padrão pode ser usado para distinguir os fósseis de nautiloides dos de amonoides. E, no seio dos amonoides, distinguir de modo expedito as principais ordens da Classe Ammonoidea: Ordem Goniatitida, Ceratitida e Ammonitida. Mas atenção, são reconhecidas mais ordens de amonoides e, também, mais tipos de linha de sutura.
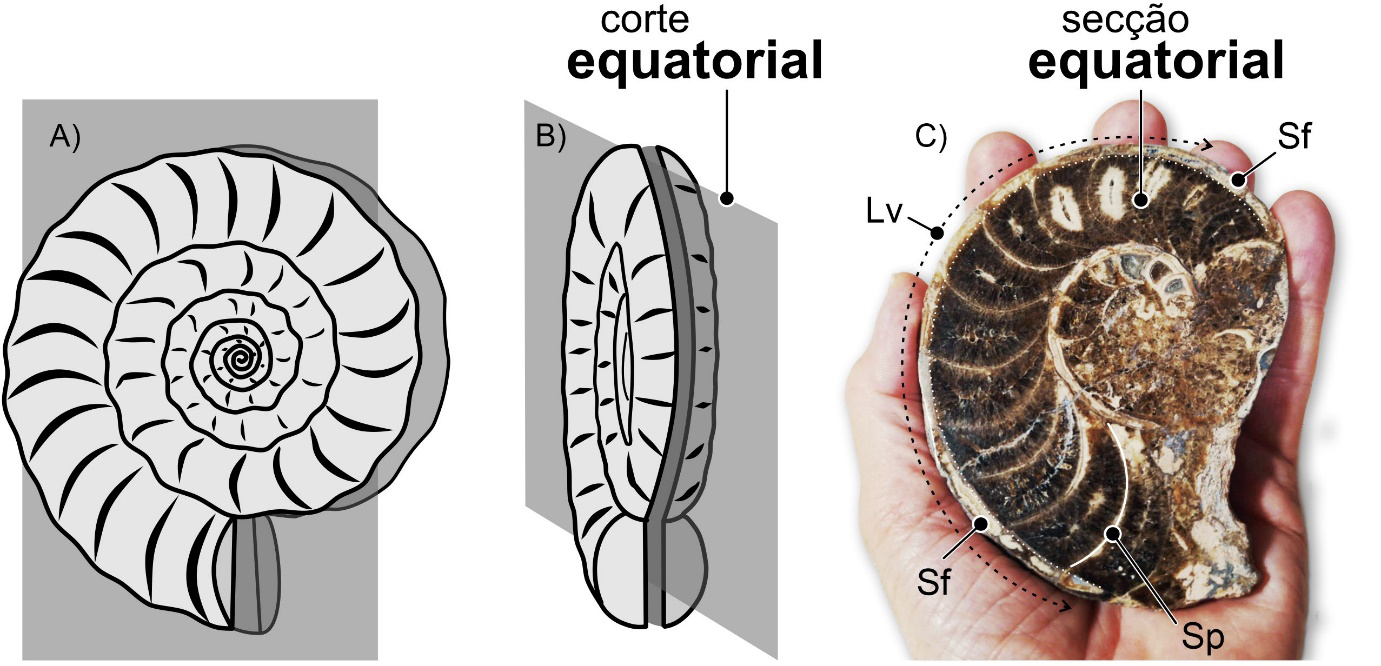
Em secção, em contexto urbano, segundo cortes equatoriais, os critérios mais fiáveis para distinguir fósseis de nautiloides nautilicónicos dos de amonoides, nomeadamente de amonites, são o carácter dos septos e o posicionamento do sifão, quando visível. Nos nautiloides, em secção longitudinal (nos ortocónicos) e equatorial (nos nautilicónicos) os septos apresentam a sua concavidade dirigida para diante, em direção ao habitáculo (FIGURA 6C e 7C). São septos ditos pró-célicos. Já nos amonoides, em secção equatorial, os septos apresentam-se, mais comummente, com a concavidade dirigida para trás, para o início do enrolamento. São septos opistocélicos (FIGURA 12C). Em secções subequatoriais ou oblíquas, nos amonoides, os septos – porque possuíam superfícies mais complicadas – podem apresentar aspetos muito variados, irregulares (e.g., FIGURA 13B). No que toca ao posicionamento do sifão, uma vez mais, em secção longitudinal e equatorial, nos nautiloides ele é – predominantemente – subcentral (FIGURA 6C e 7C). Por vezes excêntrico, desviado para o lado ventral (FIGURA 8B). Nos amonoides, mais comummente, o sifão ocupa uma posição ventral na concha (FIGURA 12C). Contudo, o sifão nos fósseis de amonoides, devido ao seu diâmetro significativamente mais reduzido e à posição junto ao bordo ventral interno da concha, não é fácil de observar. Não é, normalmente, visível em secções expostas em fachadas ou pavimentos urbanos.

Para reconhecer fósseis de nautiloides em secção, em contexto urbano, os cortes mais adequados – aqueles que geram secções mais informativas – são os cortes longitudinais, no caso das conchas ortocónicas (FIGURA 7C) e os equatoriais (FIGURA 6C)), no caso das conchas nautilicónicas. Longitudinais, são os cortes que se desenvolvem ao longo do eixo de crescimento das conchas ortocónicas. Sabemos que um corte é longitudinal quando na secção resultante é visível o sifão ao longo de todo o seu comprimento. Se isto não suceder, as secções resultaram de cortes com orientações diferentes. De cortes sublongitudinais, se se desenvolveram ao longo da concha, mas não forem coincidentes com o sifão. Ou de oblíquos, quando resultam de cortes realizados segundo diferentes ângulos em relação ao eixo da concha.
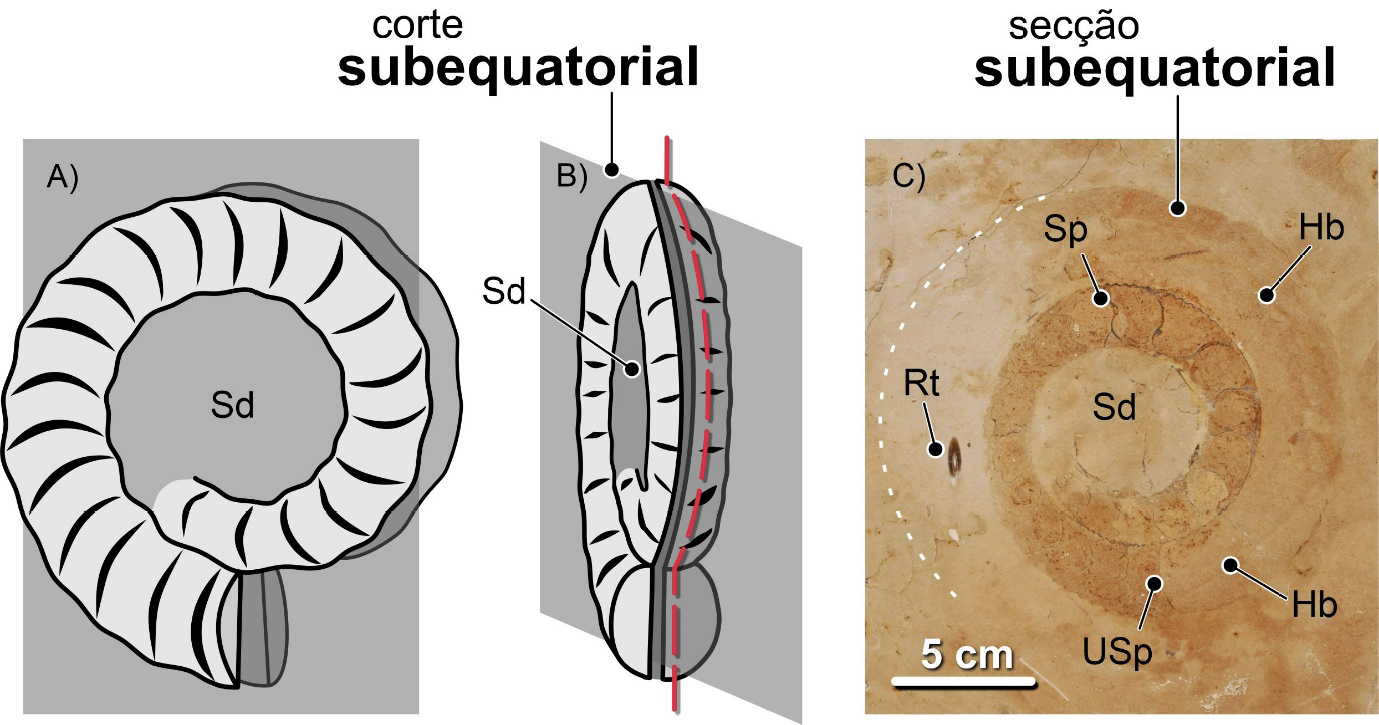
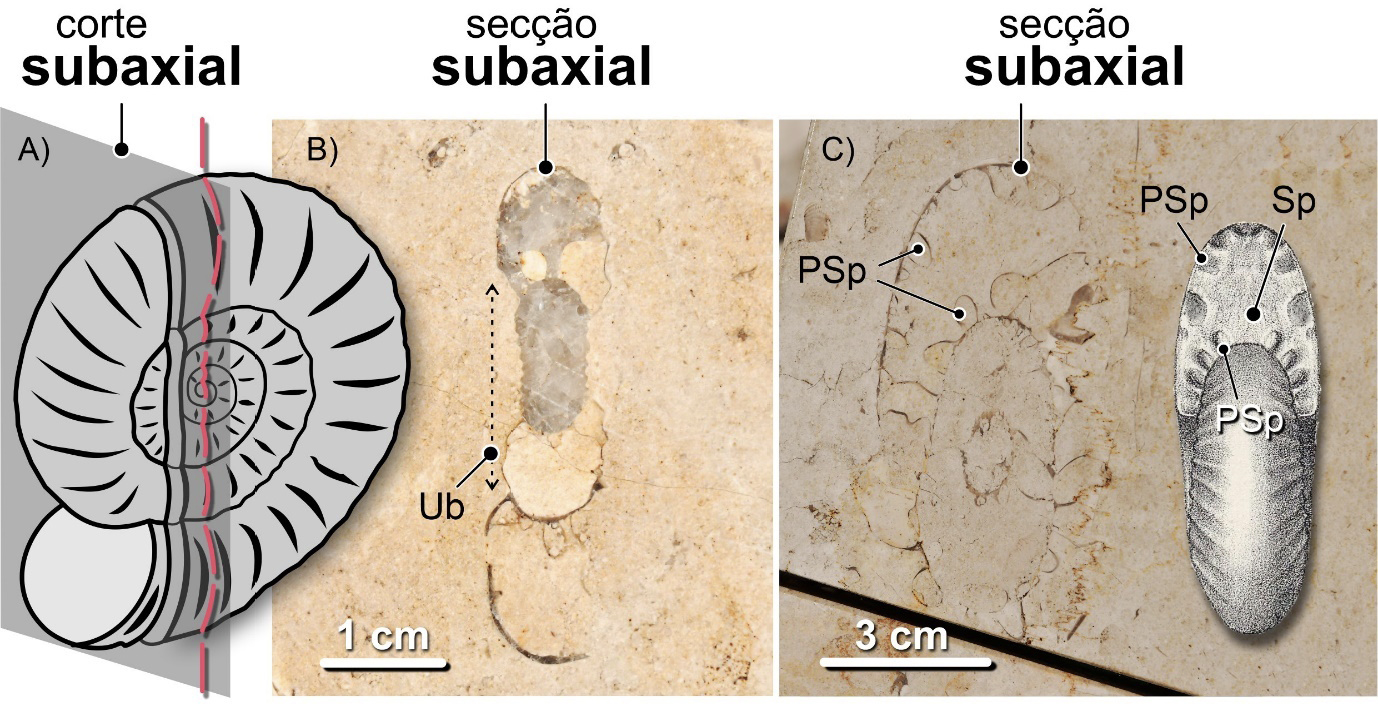
Nas conchas nautilicónicas, os cortes equatoriais são os que coincidem com o plano médio da concha, perpendiculares ao eixo de enrolamento. Numa secção equatorial perfeita, o sifão da concha será visível ao longo de toda a sua extensão. Se isso não acontecer, o corte será subequatorial – i.e., não é equatorial, mas quase, paralelo ao plano médio da concha – ou, mais frequentemente, oblíquo, segundo um angulo diferente de 90º em relação ao eixo de enrolamento. Os cortes axiais ou transversais são os que coincidem com o eixo de enrolamento da concha. Se os cortes se desenvolverem perpendicularmente ao plano equatorial da concha, mas não coincidirem com o eixo de enrolamento, serão subaxiais (FIGURA 15).
Os amonoides e os seus fósseis na cidade.
A primeira referência aos fósseis a que hoje damos o nome de amonites remonta ao naturalista romano Plínio “o Velho” (AD ?23-79), falecido em 24 de agosto de 79 em Pompeia, aquando da famosa erupção do Vesúvio. Na sua obra História Natural, Plínio refere, a dada altura: “Hammonis cornu inter sacratissimas Aethiopiae, aureo colore arietini cornus effigiem reddens, promittitur praedivina somnia repraesentare.”23. (Dizem que o corno de Amon, uma das mais sagradas [pedras] da Etiópia, com sua cor dourada [i.e., piritizada] e lembrando um chifre de carneiro, proporciona sonhos premonitórios).

É desta referência que surge o nome Amonite, de “Amon”, deus egípcio figurado com cornos espiralados de carneiro, sobretudo nas representações helenísticas de Amon-Zeus, seguido do sufixo “-ite” vulgar em elementos geológicos, em fósseis e minerais. Tal como em trilobite, biotite, belemnite, caulinite, etc. É também desta referência aos “cornos de Amon” que resulta a terminação “-ceras” comum nos nomes dos géneros de muitos amonoides e nautiloides: Dactylioceras, Harpoceras, Orthoceras, etc. “Ceras” resulta do grego antigo κέρας, kéras, corno.
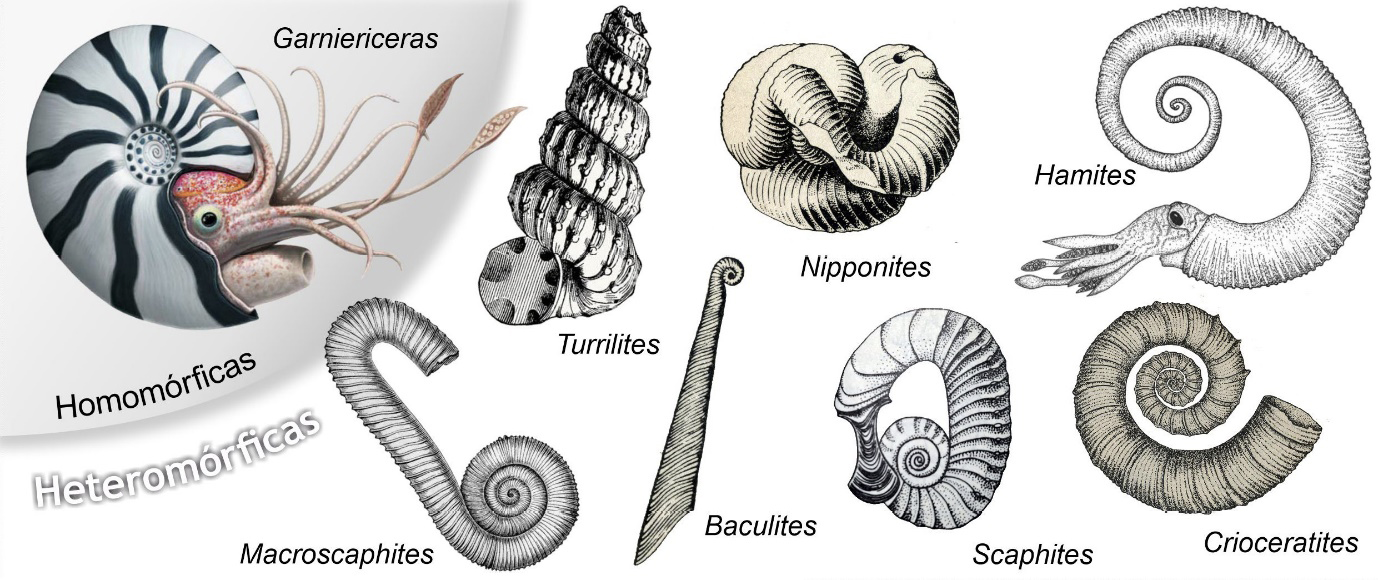
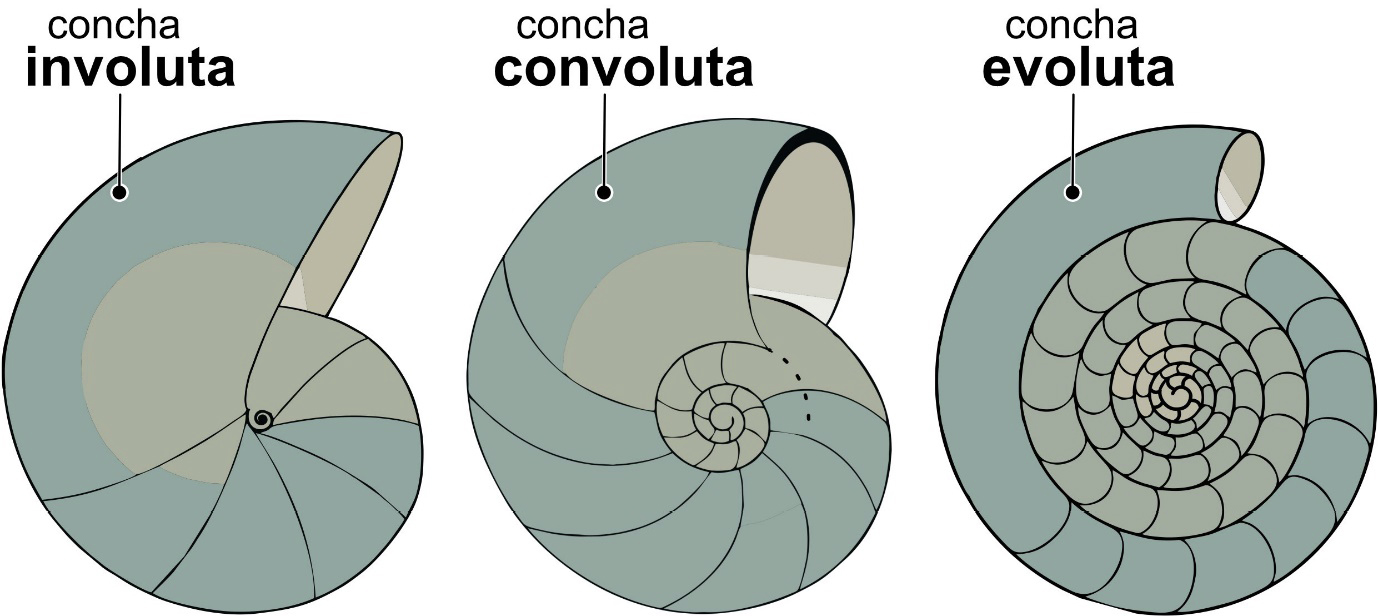
Os amonoides, ao longo dos seus 344 Ma de existência e de evolução, do Devónico até finais do Cretácico (FIGURA 2), desenvolveram conchas com formas muito variadas (FIGURA 10), bastante mais diversas que as dos nautiloides (FIGURA 3). Em termos gerais, dois tipos fundamentais de conchas de amonoides, ou melhor, de tipos de enrolamento da concha, são distinguidos: enrolamento homomórfico e heteromórfico. Quanto ao modo de enrolamento, entre as amonites homomórficas, destacamos conchas evolutas, convolutas e involutas (FIGURA 11). Há que ter em conta que estas são categorias criadas pelos humanos para lidar com a espantosa diversidade de formas de conchas de amonoides, para as catalogar e descrever. Não são categorias naturais estanques. Na natureza existe um continuum de formas, das conchas claramente involutas às tipicamente evolutas, passando pelas mais ou menos convolutas.
Assim como na arquitetura, no mundo biológico, presente e passado, a forma é a materialização – a corporização – da função. Ou seja, a morfologia dos organismos é indissociável da maneira como eles funcionam e de como se relacionam com o meio físico em que vivem, i.e., do seu modo de vida2. Diferentes tipos de conchas refletem diferentes modos de estar no ambiente, de se relacionar com o meio físico aquático em que estes organismos viviam.
As conchas com enrolamento homomórfico são as que nos são mais familiares, as que nos vêm imediatamente à mente quando pensamos em amonites. São aquelas que apresentam enrolamento planispiralado ao longo de toda a vida do animal. Nestas a espiral da concha desenvolve- se num único plano, perpendicular ao eixo de enrolamento. Os amonoides heteromórficos apresentam enrolamentos que se afastam deste tipo básico planispiralado, seja de que modo for. Por vezes, de modos extraordinários (FIGURA 10). Não é fácil entender – de imaginar! – como viviam organismos de grupos biológicos extintos há mais de 66 Ma. Organismos muito diferentes dos que conhecemos da atualidade. Contudo, existem várias hipóteses tentando relacionar a forma da concha com o modo de vida destes animais. Assim, segundo a mais popular, os amonoides homomórficos seriam organismos pelágicos nectónicos, porventura mais ágeis, nadando ativamente na coluna de água. Os heteromórficos teriam modos de vida distintos, de pelágicos mais passivos, planctónicos, até demersais, vivendo junto ao fundo27. Talvez algumas amonites heteromórficas demersais tivessem também um modo de alimentação, um tipo trófico, que se afastaria do de predadores ativos, mais comum, por exemplo, entre os cefalópodes pelágicos nectónicos. Seriam, possivelmente, segundo alguns autores, suspensívoros, alimentando-se de organismos de pequenas dimensões, de zooplâncton28.
Ao inverso do que em tempos se supôs, a diversidade de formas das amonites heteromórficas cretácicas não reflete a degenerescência do grupo, prenúncio de extinção, por se afastarem do “ideal” homomórfico mais familiar. Pelo contrário, tal como para os rudistas2, revelam uma capacidade do grupo de se diversificar, de explorar novos habitates e novas maneiras de “estar na vida”.
Em Portugal, em contexto urbano, apenas estão documentados fósseis de amonites homomórficas, planispiraladas (e.g., FIGURA 5A)). Talvez em paragens mais excitantes existam exemplos de conchas heteromórficas patentes em fachadas ou pavimentos.
Também no que toca a dimensões as conchas dos amonoides eram bastante variadas. O maior fóssil destes animais conhecido, um molde interno da concha da amonite Parapuzosia (P.) seppenradensis do Cretácico Superior da Alemanha, está exposto no Museu de História Natural da Vestefália, Westfälische Museum für Naturkunde, em Münster. Tem cerca de 1,8 m de diâmetro30. Estima-se que a concha original, completa, poderia atingir 2,5 a 3,0 m de diâmetro. Um verdadeiro gigante dos mares, capaz de rivalizar com o Kraken! Felizmente, as dimensões mais comuns destes fósseis rondam desde alguns centímetros até 20 cm de diâmetro.
Os fósseis de grandes dimensões impressionam. Sim! Mas não são práticos, nem – verdadeiramente – úteis. Em trabalho de campo, fazendo o levantamento paleontológico e estratigráfico de uma dada região, por vezes em zonas de muito difícil acesso, durante o qual se recolhem dezenas de exemplares para estudo, não é nada fácil coletar exemplares com 30 cm ou mais de diâmetro. Quantos fósseis desses se conseguiriam carregar na mochila para trazer para o laboratório, ou para o museu, tendo de caminhar vários quilómetros por montes e vales?!
Em cidade, devido às dimensões relativamente reduzidas dos materiais usados em construção, seria complicado ter evidências de amonoides do tamanho de uma amonite do género Parapuzosia. Contudo, são conhecidas ocorrências de fósseis de amonites com cerca de 50 cm de diâmetro, como se pode ver nos pavimentos da estação de Metro Reboleira, em Lisboa (FIGURA 16D).
Os fósseis urbanos destes e doutros amonoides são expostos como resultado de cortes com orientações variadas. Como resultado, à semelhança do que sucede com as conchas de nautiloides nautilicónicos, obtemos três categorias principais de secções: equatoriais (= longitudinais), as mais fáceis de reconhecer, mostrando claramente o crescimento espiralado da concha, axiais (=transversais) e oblíquas. As secções equatoriais resultam de cortes orientados ao longo do máximo diâmetro da concha, perpendicularmente ao eixo de enrolamento (FIGURA 12). As secções axiais, de cortes orientados ao longo do eixo de enrolamento dos amonoides homomórficos. As oblíquas, segundo planos posicionados entre as axiais e as equatoriais segundo ângulos variados, não sendo paralelas nem às axiais, nem às equatoriais (FIGURA 13). Quando as secções não são coincidentes com o plano de máximo diâmetro da concha, dizem-se subequatoriais (FIGURA 14). Do mesmo modo, quando resultam de cortes paralelos ao eixo de enrolamento axial, não coincidindo com ele, dizem-se subaxiais (FIGURA 15).
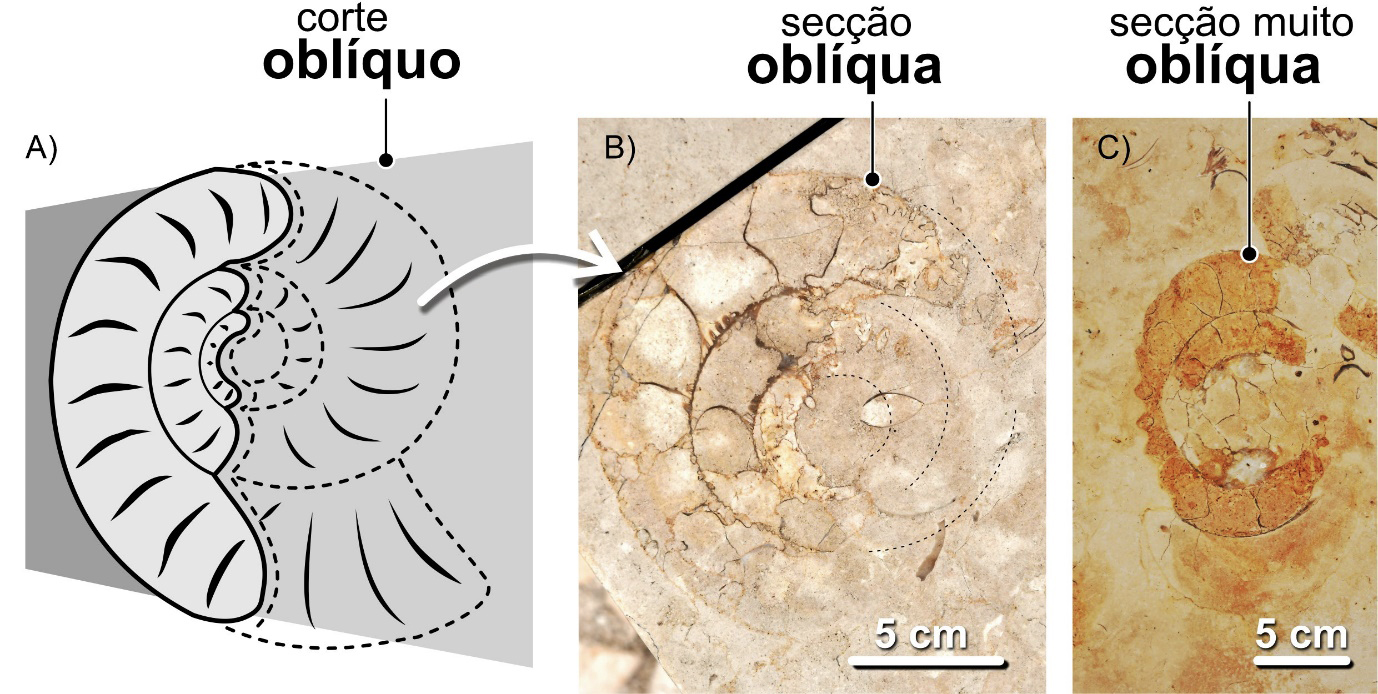
Em Portugal, uma vez que as rochas ornamentais em que estes fósseis ocorrem são, sobretudo, de idade Jurássico, encontramos vestígios de amonites, já que goniatites e ceratites, em tempos jurássicos, já não existiam (FIGURA 16). Em contexto urbano, os fósseis de amonites mais fáceis de descobrir e de identificar são os que se apresentam em secções equatoriais e subequatoriais, pois o seu aspeto espiralado é facilmente reconhecível, chamando a atenção mesmo dos mais distraídos. Distinguir e identificar fósseis seccionados segundo outras orientações poderá ser mais desafiante. Contudo, nada que imaginar as conchas destes animais a três dimensões e a prática não resolvam.
O que torna os fósseis de amonites tão úteis para datação?
Os fósseis dão-nos inúmeras informações. São ferramentas fundamentais – entre muitas outras coisas – para determinar a idade dos estratos rochosos em que ocorrem. São usados com sucesso para este fim – consistentemente – desde finais do séc. XVIII. Desde muito antes do advento do modelo de evolução por seleção natural de Darwin (1809-1882) em 1859. Muito antes de se entender a biodiversidade como uma entidade que se transforma evolutivamente ao longo do tempo. Sendo essa transformação, precisamente, o que permite que os fósseis sejam usados como padrões temporais. Ao longo do tempo surgem e extinguem-se espécies, grupos biológicos. E os grupos biológicos só surgem uma vez e, quando se extinguem, é para sempre. E isso fica registado nas sequências estratigráficas. As amonites surgiram no início do Jurássico e extinguiram- se no final do Mesozoico. Isso significa que quando encontramos fósseis de amonites, a idade de formação daqueles estratos estará contida naquele período de tempo, Jurássico-Cretácico.
Quando usamos os fósseis para estabelecer a cronologia de estratos geológicos, em conjugação, por exemplo, com o Princípio da Sobreposição de Niels Steensen (1638 -1686), mais conhecido entre nós como Nicolau Steno, é costume falar-se de “datação relativa”. Por oposição à “datação absoluta” resultante de, por exemplo, métodos radiométricos, isotópicos, aqueles que nos dão um valor em milhões de anos.
Acontece que, na linguagem corrente, quotidiana, os termos “relativo” e “absoluto” carregam uma carga subliminar que é perniciosa neste contexto. O que é relativo é visto como condicional, inconstante, como dependendo das circunstâncias. Logo, como impreciso, contingente ou fortuito. Costuma dizer-se “isso é relativo”, quando queremos retirar a algo o seu carácter indiscutível ou irrefutável. Já o que é absoluto é entendido como definitivo, preciso, irrefutável. “Ter certeza absoluta”, diz-se.
Neste caso, “relativa” alude ao facto de este método de datação se basear na posição de umas associações de fósseis em relação às outras nas sequências estratigráficas. Ou seja, uma vez estabelecida a sequência ou ordem de ocorrência das associações fossilíferas, uma vez conhecida a relação vertical entre as associações fossilíferas, quando encontramos uma delas, sabemos em que ponto da história da Terra estamos, sem necessidade de lhe atribuir uma idade em milhões de anos.
A idade em milhões de anos é-nos dada pela datação radiométrica, numérica. Contudo, este método de datação não é mais preciso, nem mais fidedigno que a datação realizada com base nos fósseis. Por vezes, é até menos fiável. Depende das circunstâncias. Ambos os métodos de datação são utilizados. Ambos têm vantagens e desvantagens. Ambos são complicados de aplicar, requerendo formação especializada.
A radioatividade foi descoberta por Henri Becquerel (1852-1908) em 1896 e a datação radiométrica foi tentada pela primeira vez por Ernest Rutherford (1871-1937) apenas em 190631. Contudo, até essa altura – desde inícios do séc. XIX – já se havia desenvolvido uma cronologia da história da Terra bastante detalhada, baseada nos fósseis e na sua datação relativa, posicional. Só não se sabia qual a duração temporal de cada divisão entretanto estabelecida.
Também no dia-a-dia usamos estes dois tipos de cronologias. Um exemplo de datação posicional (relativa): O reinado de D. Diniz teve lugar entre o de Afonso III, antes, e o de Afonso IV, depois. Já de um ponto de vista da datação numérica, diríamos: O reinado de D. Diniz decorreu de 1279 a 1325. Poderíamos pensar que a datação numérica seria mais útil. Em algumas circunstâncias, é. Contudo, por exemplo, com os resultados da datação numérica não conseguiríamos responder à questão: quem foi o pai de D. Diniz? Vantagens e desvantagens!
Assim, seria útil, neste contexto, usar terminologia que não carregasse a bagagem subjetiva acima referida. Datação posicional, em vez de datação relativa, por exemplo, e datação numérica, datação isotópica ou até radiométrica, em vez de absoluta, seriam expressões mais úteis.
A utilidade biostratigráfica dos fósseis, isto é, o seu préstimo como ferramentas de datação, está associada a uma série de características dos grupos biológicos do passado que lhes deram origem que, por sua vez, determinam um conjunto de particularidades dos próprios fósseis. Neste contexto, aplica-se a regra informal EGAS. Não, não é uma homenagem ao famoso Egas da Rua Sésamo. A ideia é que quanto melhor os fósseis cumprirem a regra EGAS, mais úteis serão biostratigraficamente. EGAS é o acrónimo de Estratigrafia, Geografia, Abundância e Sistemática, i.e., facilidade de identificação do grupo biológico correspondente.
Estratigrafia. Quanto mais curta a expressão estratigráfica dos fósseis de um determinado grupo biológico, mais úteis serão para identificar um intervalo de tempo passado em concreto e, consequentemente, melhores serão como indicadores de idade, como elementos de datação.
As amonites foram um grupo biológico muito dinâmico, evolutivamente falando. Espécies novas estavam constantemente a surgir e a extinguir-se, sendo substituídas por outras, de um modo rápido à escala do tempo geológico. Resultado, os seus fósseis, nas sequências de estratos jurássicos e cretácicos de hoje, têm expressão estratigráfica curta. Têm extensão vertical reduzida nas sequências de camadas geológicas, de estratos. São bons indicadores estratigráficos.
Geografia. Quanto mais ampla a distribuição geográfica dos fósseis de um determinado grupo biológico, mais úteis serão para correlacionar, i.e., para comparar temporalmente os estratos geológicos das diferentes regiões onde ocorrem.
Os cefalópodes são organismos essencialmente marinhos. Os seus fósseis ocorrem em rochas sedimentares de génese marinha. No caso dos amonoides, eram maioritariamente organismos pelágicos nectónicos, i.e., nadadores em águas oceânicas. Por isso, estes grupos biológicos gozavam de ampla distribuição geográfica. Consequentemente, hoje em dia, os seus vestígios fossilizados também. Ou seja, podemos encontrar fósseis das mesmas amonites jurássicas aqui em Portugal e, por exemplo, no sul de França, na Grécia e no norte de África. Isso é bom, pois permite atribuir a mesma idade a todas essas ocorrências por via do Princípio da Identidade Paleontológica. Associações de fósseis iguais, idade igual.
Além disso, as amonites eram organismos pelágicos nectónicos, nadadores, não estavam dependentes das condições do fundo. Por exemplo, não eram condicionadas pelas características dos sedimentos. Por isso, quando morriam, as suas conchas acabavam enterradas em sedimentos muito diversos. Resultado, os seus fósseis podem ocorrer em rochas variadas, de argilitos a calcários, passando por arenitos, para referir apenas as mais comuns. Isto é, podem ocorrer em fácies variadas. Ou seja, os fósseis dos cefalópodes pelágicos são facialmente independentes. Esta independência das fácies, i.e., do tipo de rocha associado a um ambiente de deposição, é uma mais-valia para a utilidade biostratigráfica dos fósseis dos cefalópodes, pois significa que a probabilidade de encontrarmos os seus fósseis é mais elevada.
Abundância. Quanto mais fósseis de um determinado grupo biológico existirem, mais fáceis serão de encontrar e, consequentemente, mais fáceis de usar para datação.
Para se usar os fósseis para datar é necessário encontrá-los. Como já referido no capítulo “Fossilização. Fenómeno raro?”, acima, a existência de concha resistente, por exemplo, favorece o registo fossilífero dos animais que a possuem. Ora, as amonites possuíam concha. E eram organismos muito importantes nos ecossistemas marinhos da altura. Consequentemente, existiam em grande quantidade. Logo, hoje, existem muitos fósseis de cada espécie de amonite.
Por outro lado, as amonites eram animais marinhos. Em ambientes aquáticos, nomeadamente nos marinhos, predominam os fenómenos de sedimentação, de acumulação de sedimento. Havendo acumulação de sedimento, é mais fácil enterrar restos orgânicos. Mais restos enterrados, mais probabilidade de haver fósseis. Pelo contrário, em ambientes terrestres – onde viviam os dinossáurios mesozoicos, por exemplo – predominam os fenómenos de erosão. A probabilidade de destruição dos vestígios orgânicos é maior, a de preservação, menor.
Resultado de tudo isto? Há muitos mais fósseis de conchas de amonites jurássicas que de dinossáurios da mesma idade. Esse é um grande ponto a favor das amonites no que toca ao seu uso para datação.
Sistemática. Quanto mais elementos morfológicos diagnósticos os fósseis de um determinado grupo biológico exibirem, mais fácil se tornará identificá-los e, consequentemente, mais eficazes serão para datar os estratos em que ocorrem.
Para se poder usar os fósseis para datar é necessário identificá-los de modo tão preciso quanto possível. Saber que encontrámos um fóssil de amonite, da Ordem Ammonitida, permite dizer que a idade dos estratos onde ocorre está contida no intervalo Jurássico a Cretácico. Mas se identificarmos a espécie, por exemplo, Dactylioceras polymorphum, saberemos as camadas onde ocorre correspondem à Zona Polymorphum32, a mais antiga do Andar Toarciano do Sistema Jurássico Inferior, uma datação posicional muito mais detalhada. O intervalo estratigráfico Jurássico – Cretácico representa um período de tempo de cerca de 135 Ma. Já a zona mais antiga do Andar Toarciano corresponde a um lapso de apenas cerca de 1 Ma. Um intervalo de tempo muito mais curto!
Para identificar detalhadamente um fóssil é fundamental que ele preserve aspetos variados da morfologia do organismo que permitam essa diferenciação, essa discriminação entre os vestígios de organismos das diversas espécies. As conchas das amonites apresentavam formas muito variadas: heteromórficas e homomórficas, evolutas, convolutas, involutas (FIGURA 10, 11). A superfície das suas conchas – ao contrário do que sucedia, e.g., com as conchas dos nautiloides – era, frequentemente, muito ornamentada. Isto é, possuíam relevos muito variados. Apresentavam costilhas radiais simples, bifurcadas, trifurcadas, tubérculos, espinhos, carenas e sulcos ventrais, etc. Todos estes aspetos permitem estabelecer uma classificação detalhada das amonites. Se as conchas fossem todas iguais, com o mesmo tipo de enrolamento, a mesma forma e lisas, i.e., sem ornamentação, ou se aquilo que as distinguia não ficasse preservado – a cor, por exemplo –, não seria possível individualizar as diferentes espécies. Consequentemente, não seriam tão úteis para datação.
Resumindo, quanto melhor um determinado grupo de fósseis cumprir as quatro características acima referidas – expressão estratigráfica curta, distribuição geográfica ampla, existirem em quantidade e apresentarem elementos variados de identificação – mas úteis serão para datação!
É comum, abordando este tema, usar-se a dicotomia simplista e algo antiquada Fósseis de Fácies versus Fósseis de Idade. Contudo, há fósseis – como os das amonites – que são excelentes para datação de estratos de idade Jurássico-Cretácico e, também, para interpretação paleoambiental, pois são evidências de ambientes marinhos típicos. Se virmos o mundo como uma coleção de “caixinhas”, em qual delas colocaremos os fósseis de amonites? Teríamos de os colocar ao mesmo tempo nas duas. Lá se vai a dicotomia.
Do acima exposto resulta que não faz sentido criar caixinhas como Fósseis de Fácies e Fósseis de Idade. O mundo não é a preto e branco. E os fósseis também não! Pelo contrário, o mundo é feito de nuances. Seria mais vantajoso abordar-se a utilidade dos fósseis como elementos de datação e como instrumentos de interpretação paleoambiental, paleoecológico, em geral, sem os colocar em gavetinhas. Isto porque todos os fósseis encerram, para além de muitas outras categorias de informação, algum tipo de referência cronológica, uma vez que os grupos biológicos que estiveram na sua origem existiram num certo intervalo de tempo, e de informação paleoambiental, pois viveram num determinado ambiente.
E, claro, não perder de vista todas as outras informações que os fósseis nos dão sobre, e.g., evolução, fossilização, biogeografia, tafonomia, paleogeografia, etc.
Os coleoides e os seus fósseis na cidade.
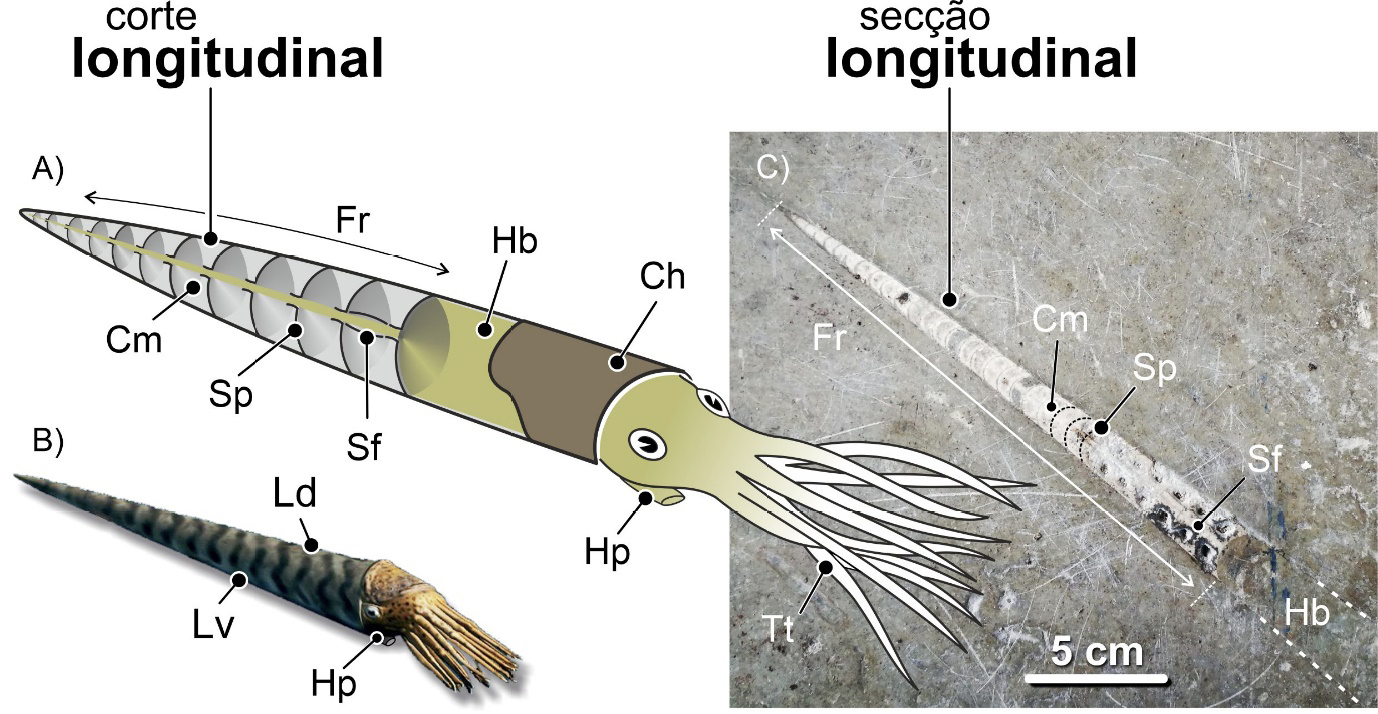
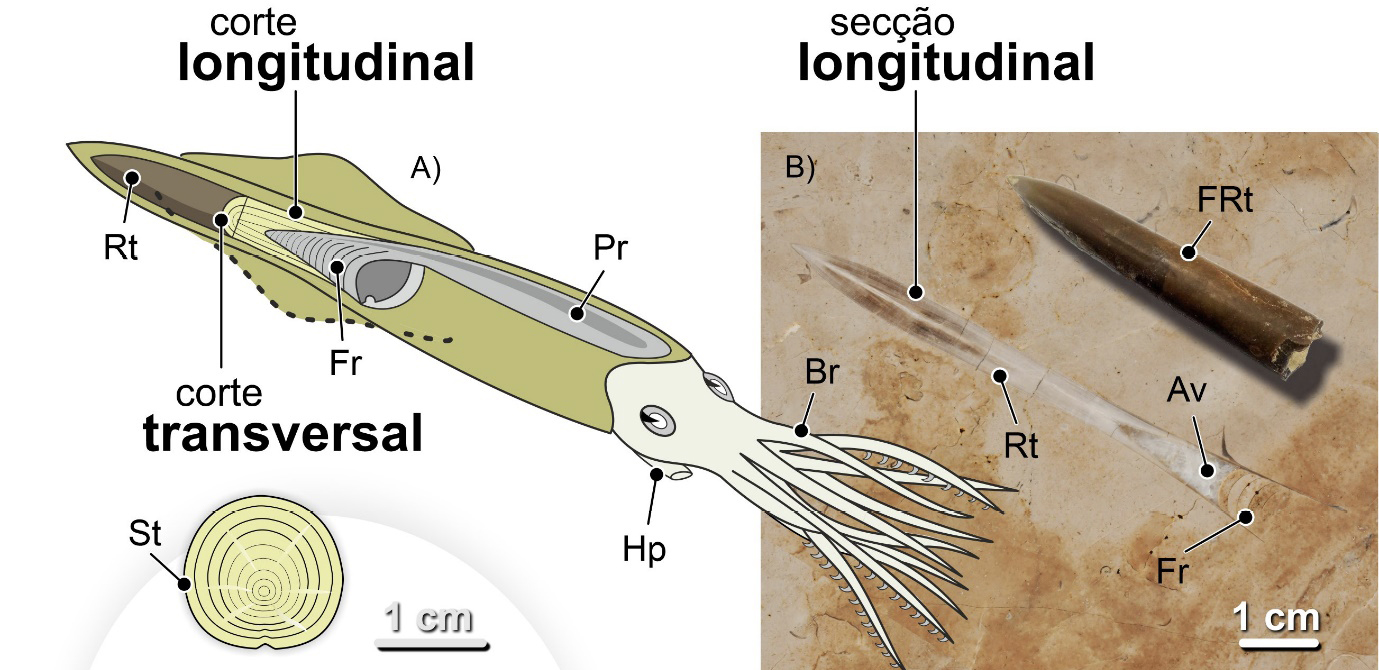
Os coleoides da atualidade, chocos, lulas e polvos, como vimos, não possuem elementos esqueléticos que favoreçam a sua fossilização. Existem fósseis destes organismos, mas são bastante invulgares. Assim sendo, não é de espantar que os fósseis mais comuns de coleoides, nomeadamente em contexto urbano, sejam os dos rostros da concha interna das belemnites, um grupo extinto de cefalópodes. A sua forma alongada, lembrando um projétil, ou uma ponta de dardo, quando vista em secção longitudinal, é inconfundível (FIGURA 17).
Essa forma de projétil, de resto, valeu o nome ao grupo Belemnoidea, do grego antigo βέλεμνον, bélemnon, significando dardo, projétil. O sufixo -oidea, ou -oide, em português, também de origem grega, significa “com forma, com aparência de”.
Este aspeto de projétil não lhes valeu apenas o nome. Também originou algumas lendas associadas à sua ocorrência em afloramentos naturais. Por exemplo, em Inglaterra, à ocorrência comum de inúmeros de fósseis de rostros de belemnites no mesmo local, por vezes às centenas, é dado o nome de “belemnite battlefields”33, campos de batalha de belemnites. A expressão teve origem no famoso paleontólogo de cefalópodes alemão Friedrich August von Quenstedt (1809-1889), por – nas palavras dele – parecer que naqueles locais “se terá travado uma verdadeira batalha”34 da qual apenas restam as balas espalhadas.
Também em Portugal há histórias semelhantes. Em Peniche, em tempos, pelo menos até à década de 1970, o troço de costa norte da península entre a Ponta do Trovão e a Praia do Portinho na Areia do Norte era conhecido entre os pescadores locais como o sítio da antiga fábrica de munições. “As balas ainda por lá andam pelo chão”, diziam, explicando o insólito nome.
As belemnites, em termos gerais, tinham aparência próxima da das lulas atuais. Era animais de corpo alongado, em forma de torpedo, possivelmente com barbatanas laterais alongadas e possuindo saco de tinta35. Sabemos isto pois existem alguns fósseis excecionais destes animais em que é discernível – ainda que de modo vago – o contorno do seu corpo. Por essa mesma razão, sabe-se também que – ao contrário das lulas que possuem oito braços e dois tentáculos – possuíam apenas 10 braços de igual tamanho munidos de ganchos córneos, quitinosos.
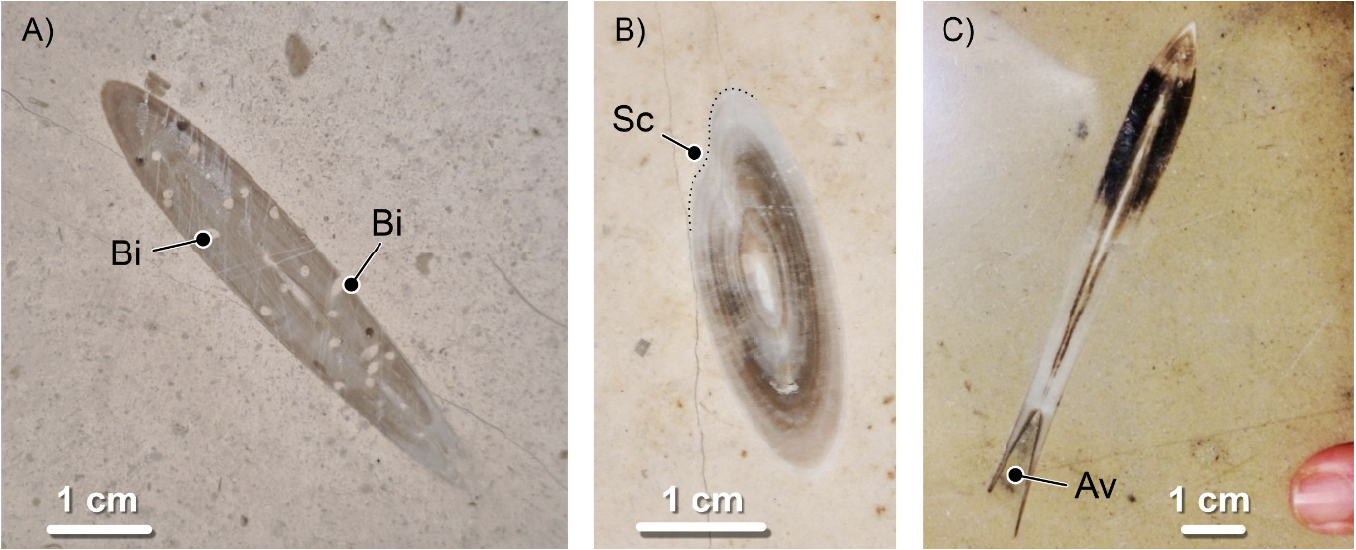
A concha interna das belemnites é normalmente dividida em três zonas ou elementos constituintes principais. Do extremo distal para o proximal, isto é, de trás para a frente do molusco: rosto, fragmocone e pro-óstraco (FIGURA 17). Estes elementos tinham composições mineralógicas e comportamentos mecânicos distintos. O rostro era maciço, constituído pelo mineral calcite, e bastante resistente. O fragmocone e o pró-óstraco, significativamente mais delicados. Além disso, eram constituídos por aragonite, um mineral que, apesar de ter a mesma constituição química da calcite, tem propriedades distintas. A aragonite biogénica, quando enterrada nas camadas mais superficiais da crosta terrestre após a morte dos organismos, é menos resistente quimicamente, mais solúvel, sendo mais facilmente eliminada. Assim, há muitos fósseis de rostros e muito poucos em que toda a concha tenha sido preservada.
Os fósseis de belemnites, em contexto natural, no nosso país, poderão ser encontrados em rochas de idades Jurássico Inferior, na região de São Pedro de Moel, a Cretácico Inferior, em Cascais e no Cabo Espichel36. Já em contexto urbano, uma vez mais, encontraremos sobretudo fósseis de belemnites jurássicas, pois as rochas ornamentais em que normalmente ocorrem são dessa idade. Não são conhecidos, até ao momento, fósseis de belemnites, por exemplo, do Liós cretácico da região de Lisboa tão abundante em várias cidades nacionais.
Sendo os rostros os elementos mais resistentes do esqueleto interno destes animais, serão estes que encontraremos mais frequentemente fossilizados. São sobretudo fáceis de reconhecer quando se apresentam sob a forma de secções longitudinais (FIGURA 17).
Os rostros apresentam dois padrões microstruturais: um concêntrico resultante dos anéis de crescimento dispostos homocentricamente em redor do eixo central do rostro e, sobreposto a este, um radial expressando o desenvolvimento fibroso dos cristais de calcite. O padrão concêntrico reflete o crescimento do rostro que, por sua vez, resulta do crescimento do organismo. Já o radial manifesta o desenvolvimento dos cristais de calcite que constituem o rostro. Acontece que, em contexto urbano, em fachadas e pavimentos, estando os fósseis polidos, apenas o padrão concêntrico é discernível, mais evidente em cortes transversais e oblíquos (FIGURA 18B). O extremo proximal do rostro é oco. A essa cavidade é dado o nome de alvéolo (FIGURA 18C e 19). O alvéolo alojava o fragmocone, muito reduzido, da concha do animal e pode ser visto em cortes longitudinais bem preservados (FIGURA 17).

O que fazer com estes fósseis?
Em contexto urbano, a geodiversidade pode ocorrer in situ, em afloramentos naturais no seio do tecido urbano, ou – mais frequentemente, de modo mais evidente – ex situ38, sob a forma de geomateriais de construção trazidos para a cidade para a edificar. Neste texto focam-se apenas elementos de geodiversidade ex situ na cidade.
Estes aspetos podem ser abordados de múltiplas maneiras, tantas quantos os valores que, em geral, atribuímos às manifestações da natureza geológica. Podem ser focados do ponto de vista económico, patrimonial, estético, lúdico, cultural43, etc. e, claro, científico e educativo.
Quanto aos fósseis de cefalópodes, em concreto, desde a simples fruição estética das suas formas espiraladas – ou outras – à curiosidade de os identificar em edifícios, passando pelo aspeto lúdico de “caça ao tesouro”, de procura desses vestígios na cidade, as possibilidades são bastante variadas. Contudo, talvez o modo mais óbvio de valorizar os fósseis urbanos seja usá-los no ensino e em atividades de divulgação da Paleontologia, da Geologia e da geodiversidade. O valor educativo da geodiversidade em contexto citadino foi já amplamente focada noutras publicações1, 2, 39, 40, 41 pelo que não será discutido em profundidade aqui.
No ensino, quer abordando temas geológicos, quer biológicos, os fósseis urbanos são um recurso vantajoso, excitante e muito acessível. Permitem abordar e exemplificar, em concreto, a extinção de grupos biológicos e a sua relação com o uso dos fósseis para investigar a história e a cronologia do planeta, ou seja, possibilitam abordar as rochas como arquivos que registam a história da Terra. Proporcionam o desenvolvimento em contexto urbano dos temas da interpretação paleoambiental e da ligação entre a biodiversidade, envolvendo organismos de grupos biológicos do passado, e a geodiversidade, representada pelos seus fósseis. São também um excelente ponto de partida para realçar o imperativo ético – geoético – de proteger a geodiversidade e de conservar a natureza.
É fácil constatar que na cidade de Lisboa, como em muitas outras cidades portuguesas, os fósseis de cefalópodes não são tão comuns quanto os de rudistas no Liós cretácico. Mas, mesmo assim, não são invulgares, podendo ser encontrados associados a muitas outras rochas ornamentais, sobretudo de idade Jurássico. Assim, em concreto, que abordagens se poderiam tomar? Que atividades se poderiam desenvolver? Eis aqui, de seguida, algumas possibilidades.
Por exemplo, comparar a morfologia das conchas de amonites patentes em secções equatoriais com as de Nautilus existentes da atualidade e depreender o modo de vida dos cefalópodes extintos e, consequentemente, em que paleoambientes viveriam. Neste contexto lançar-se- -ia mão do Princípio do Uniformitarismo Morfofuncional expresso – em termos gerais – como “estruturas corporais semelhantes implicam modos de vida similares”. Também o Princípio do Uniformitarismo Taxonómico poderia ser invocado, “organismos do passado de grupos biológicos extintos teriam exigências ecológicas similares às dos seus parentes mais próximos atuais”. Amonites e Nautilus possuem conchas externas tabicadas estruturadas de modo similar, isso implica que funcionariam do mesmo modo, logo as amonites homomórficas – à semelhança dos Nautilus na atualidade – seriam organismos marinhos, pelágicos nectónicos, seriam organismos nadadores ativos. E se eram pelágicos nectónicos, viviam no oceano. Este exercício permitiria abordar de modo prático o tema da interpretação paleoambiental com base no conteúdo fossilíferos das rochas.
Outra atividade. Observar, registar – i.e., descrever, desenhar ou fotografar – e caracterizar os fósseis das conchas espiraladas de amonites (evolutas, com muitas voltas, apresentando crescimento geralmente lento) em secção equatorial e procurar animais cefalópodes exatamente iguais na atualidade. Conclusão, não existem cefalópodes com características exatamente idênticas na atualidade. As amonites são um grupo extinto! Este exercício poderia ser associado a um tema focando a Terra como um planeta em mudança, quer biologicamente, por via da evolução biológica, quer geologicamente.
À semelhança da atividade anterior, procurar fósseis do esqueleto interno de belemnites e procurar animais cefalópodes com estruturas semelhantes na atualidade. Mais uma vez, não existem cefalópodes com conchas internas daquele tipo na atualidade. As belemnites são um grupo extinto. Este poderia ser um tema focando a vida e os seres vivos, a diversidade na biosfera, quer atual, que na do passado geológico da Terra.

Os fósseis de cefalópodes urbanos poderiam também ser empregues em exercícios visando o desenvolvimento da inteligência espacial, a capacidade de raciocínio tridimensional, contribuindo para a literacia científica e artística. Por exemplo, a partir de várias secções de fósseis de cefalópodes, imaginar e reconstituir graficamente a morfologia original da concha, bem como determinar a orientação dos cortes que as originaram. Esta atividade poderia ser desenvolvida interdisciplinarmente em conjugação com temas de Geometria Descritiva42 ou Educação Visual e Tecnológica com vista ao “desenvolvimento das capacidades de ver, perceber, organizar e catalogar o espaço envolvente”.
Em suma, como anteriormente salientado2, uma das muitas vantagens de empregar os fósseis urbanos em atividades pedagógicas, nos mais variados níveis de ensino, é que para usufruir deles não é necessário equipamento sofisticado, delicado ou oneroso (FIGURA 20). Curiosidade para os procurar, um borrifador com água para os avivar em pavimentos e fachadas e uma câmara fotográfica — ou um qualquer telemóvel — para os registar é quanto basta.
In Memoriam.
Irina Aleksandrovna Mikhailova (Ирина Александровна Михайлова, 1929-2022), paleontóloga, estudiosa de amonites cretácicas internacionalmente reconhecida e saudosa Professora na Cátedra de Paleontologia da Faculdade de Geologia da Universidade Estatal de Moscovo, M.V. Lomonossov. CMS, diplomado em Geologia com especialização em Paleontologia pela UEM em 1986, foi seu discípulo direto na cadeira de Paleozoologia de Invertebrados e seu orientando de tese de licenciatura, devendo-lhe a parte de leoa da sua formação paleontológica e recordando-a sempre com enorme respeito científico, imenso carinho e extrema saudade.
Materiais de apoio a atividades pedagógicas e lúdicas de Paleontologia urbana.
Sobre fósseis urbanos de Rudistas.
Sobre fósseis urbanos de Cefalópodes.
Origem das imagens/ ilustrações.
FIGURA 4 C) — Imagem de fóssil de foraminífero Globigerina adaptada de H. Grobe em Wikipedia Commons.
{kind=link}
FIGURA 4 B) — Imagem de fóssil de Archaeopteryx adaptada de H. Raab em Wikipedia Commons.
.jpg){kind=link}
FIGURA 5 C) — Reconstituição do animal belemnite adaptada de G. Jatta em Cefalopodi viventi nel Golfo di Napoli, R. Friedländer & Sohn, Berlin. 1896.
FIGURA 6 A) — Ilustração de Nautilus adaptada de E. Tutynina em Wikipedia Commons.
{kind=link}
FIGURA 7 B) — Ilustração de Orthoceras adaptada de N. Tamura em Wikipedia Commons.
{kind=link}
FIGURA 10 — Ilustração de Hamites adaptada de N. Monks, em Wikipedia Commons.
{kind=link}
FIGURA 18 C) — Imagem de fóssil de belemnite de Isabel Coelho, Professora Adjunta ISEL, Instituto Superior de Engenharia de Lisboa. À colega, o muito obrigado dos autores pela simpática cedência da imagem.
Referências
- 1 SILVA, C. M. & PEREIRA, S., A geodiversidade urbana como recurso educativo, Rev. Ciência Elem., 9(3): 51. 2021.
- 2 SILVA, C. M. & PEREIRA, S., Breve guia de Paleontologia urbana: Fósseis de Rudistas, Rev. Ciência Elem., 11(4): 43. 2023.
- 3 COSTA, M. J., Estuário do Tejo: onde o rio encontra o mar. Editora Afrontamento. 2021.
- 4 O’SHEA, S. & BOLSTAD, K., Giant Squid and Colossal Squid Fact Sheet, The Octopus News Magazine Online, TONMO. 2019.
- 5 CARTRON, L. et al., The “prawn-in-the-tube” procedure: what do cuttlefish learn and memorize?, Behavioural Brain Research, 240, 29- 32. 2013.
- 6 CUVIER, G., Second Mémoire sur l’organisation et les rapports des animaux à sang blanc, dans lequel on traite de la structure des Mollusques et de leur division en ordre, lu à la société d’Histoire Naturelle de Paris, le 11 prairial an troisième [30 May 1795], Magazin Encyclopédique, ou Journal des Sciences, des Lettres et des Arts [1. année], 2: 433-449.
- 7 LANDING, E. et al., Proposed Early Cambrian cephalopods are chimaeras, the oldest known cephalopods are 30 m.y. younger, Commun. Biol., 6, 32. 2023.
- 8 KRÖGER, B. et al., Cephalopod origin and evolution: a congruent picture emerging from fossils, development and molecules: extant cephalopods are younger than previously realised and were under major selection to become agile, shellless predators, BioEssays, 33(8), 602-613. 2011.
- 9 FREY, R. C., Middle and Upper Ordovician nautiloid cephalopods of the Cincinnati Arch region of Kentucky, Indiana, and Ohio. 1066, US Gov. Printing Office. 1995.
- 10 JAIN, S., Fundamentals of Invertebrate Palaeontology, Springer India. 2017.
- 11 KLUG, C. et al., Anatomy and evolution of the first Coleoidea in the Carboniferous. Communications Biology, 2(1), 280. 2019.
- 12 KRUTA, I. et al., Palaeontology, 59(6), 767-773. 2016.
- 13 STEUBER, T. et al., Catastrophic extinction of Caribbean rudist bivalves at the Cretaceous–Tertiary boundary, Geology, 30: 999-1002. 2002.
- 14 SILVA, C. M., E se os dinossáurios tivessem sido exterminados por um vírus? Al-Madan, II sér., 23: 8-10. 2020.
- 15 SILVA, C. M. & COSTA, A. M., Da origem da Terra à ocupação humana: Uma história geológica em Alvalade, Santiago do Cacém. In Memórias da Terra, das Águas e dos Povos, Deus, M. (Coor.), CM Santiago do Cacém, pp. 23-36. 2021.
- 16 BARSKOV, I. S., Silurian and Devonian Nautiloids from Southern Ferghana. Byulletin Moskovskogo Obshchevestva Ispytatelei Prirody, Otdel Geologicheskii, ????????? ??????????? ????????. ??????????? ???????, ????? ?????????????, 35(4): 153-154. 1960.
- 17 HISTON, K. & MESSNER, F., A Systematic Study of upper Silurian (Ludfordian) Nautiloid Cephalopods from the Eggenfeld Section (Graz Palaeozoic, Styria, Austria). Jahrbuch der Geologischen Bundesanstalt, 160(1-4): 131-175. 2020.
- 18 MOURA, A. C. & CARVALHO, C., Mármores e calcários ornamentais de Portugal. Gestão de Artes Gráficas, SA, Amadora. 2007.
- 19 KRÖGER, B., Revision of Middle Ordovician orthoceratacean nautiloids from Baltoscandia, Acta Palaeontologica Polonica, 49 (1): 57–74. 2004.
- 20 EKDALE, A. A. & BROMLEY, R. G., Paleoethologic interpretation of complex Thalassinoides in shallow-marine limestones, Lower Ordovician, southern Sweden. Palaeogeography, Palaeoclimatology, Palaeoecology, 192(1-4), 221-227. 2003.
- 21 KRÖGER, B. et al., Early orthoceratoid cephalopods from the Argentine Precordillera (lower-middle Ordovician). Journal of Paleontol, 81(6), 1266-1283. 2007.
- 22 BELAÚSTEGUI, Z. & GIBERT, J. M., Icnofábrica de Cylindrichnus en el Mioceno de la costa de Tarragona (Cataluña, España). Paleolusitana, 1: 97-104. 2009.
- 23 EICHHOLZ, D. E., Pliny, Natural History. With an English translation. Vol. X (Libri XXXVI-CCCVII). Loeb Classical Library, Harvard Univ. Press & William Heinemann Lda., Cambridge, Ma. 1961.
- 24 MIRONENKO, A. A., The soft-tissue attachment scars in Late Jurassic ammonites from Central Russia. Acta Palaeont. Polonica, 60(4): 981-1000. 2015.
- 25 BONDARENKO, O. B. & MIKHAILOVA, I. A., Breve guia de identificação de invertebrados fósseis, Nedra, Moscovo, 2ª ed., 536 pp. 1984.
- 26 MIKHAILOVA, I. A., Sistemática e filogenia de amonoides cretácicos, Nauka, Moscovo, 280 pp. 1983.
- 27 WRIGHT, J. K., Ammonites. Fossils Explained, 63, Geology Today, 28(5), 186-191. 2012.
- 28 LANDMAN, N. H. et al., Mode of life and habitat of scaphitid ammonites, Geobios, 45(1): 87-98. 2012.
- 29 HENDRICKS, J. R., Digital Atlas of Ancient Life, Class Cephalopoda. 2019.
- 30 IFRIM, C. et al., Ontogeny, evolution and palaeogeographic distribution of the world’s largest ammonite Parapuzosia (P.) seppenradensis (Landois, 1895). Plos one, 16(11), e0258510. 2021.
- 31 RUTHERFORD, E., Radioactive Transformations, Archibald Constable and Co. Ltd., London, 287 pp. 1906.
- 32 ROCHA, R. C. B. et al., Base of the Toarcian stage of the lower Jurassic defined by the global boundary stratotype section and point (GSSP) at the Peniche section (Portugal), Episodes, Journal of International Geoscience, 39(3): 460-481. 2016.
- 33 DOYLE, P. & MACDONALD, D. I., Belemnite battlefields. Lethaia, 26(1), 65-80. 1993.
- 34 QUENSTEDT, F. A., Bemerkungen zum Geologischen Bilde Schwabens. In Sonst und Jetzt. Populirre Vortrbge iiber Geologie, 48-57. Laupp, Tiibingen. Sonst und Jetzt, Populäre Vorträge u?ber Geologie, p. 50-51. 1856.
- 35 HOFFMANN, R. & STEVENS, K., The palaeobiology of belemnites–foundation for the interpretation of rostrum geochemistry, Biological Reviews, 95(1), 94-123. 2020.
- 36 RITA, P. & DUARTE, L. V., Registo Estratigráfico das Belemnites em Portugal, Tylostoma, 3, 60-63. 2024.
- 37 DECKER, S. H. et al., Boring bryozoans: an investigation into the endolithic bryozoan family Penetrantiidae, Organisms Diversity & Evolution, 23, 743-785. 2023.
- 38 BRILHA, J., Inventory and quantitative assessment of geosites and geodiversity sites: a review, Geoheritage, 8(2), 119-134. 2016.
- 39 PEREIRA, H. & LOPES, F., Histórias gravadas nas rochas. Guia de campo: À descoberta das rochas e fósseis em Loulé, Câmara Municipal de Loulé. 2019.
- 40 PEREIRA. H. & LOPES, F., Geodiversity in our school’s walls and pavements. IV Encontro Internacional Casa das Ciências, Livro de Resumos, pp. 89-90. 2017.
- 41 SILVA, C. M., Fossils, Smartphones, Geodiversity, Internet, and Outdoor Activities: A Technological Geoeducational Bundle, Geoscience Education Indoor and Outdoor, Science Education, Springer Verlag, pp. 133-156. 2016.
- 42 Geometria Descritiva A. Documentos curriculares de referência, Aprendizagens Essenciais - Secundário I Geometria Descritiva A 10.º ano e 11.º ano de escolaridade.
- 43 SILVA, C. M., Urban geodiversity and decorative arts: the curious case of the “Rudist Tiles” of Lisbon (Portugal), Geoheritage, 11, 151-163. 2019.
Este artigo já foi visualizado 1019 vezes.