


microRNAs
📧 , 📧 , 📧
- * DCV/ Universidade de Coimbra
- ɫ DCV/ Universidade de Coimbra
- ‡ DCV/ Universidade de Coimbra
Referência Cordeiro, D., Correia, S., Canhoto, J., (2021) microRNAs, Rev. Ciência Elem., V9(2):030
DOI http://doi.org/10.24927/rce2021.030
Palavras-chave expressão genética, melhoramento de plantas, RNAses, regulação pós-transcripcional, silenciamento genético
Resumo
O avanço nas tecnologias de sequenciação e caracterização do RNA tem permitido revelar mecanismos essenciais na biologia dos organismos até recentemente desconhecidos. O transcritoma das células engloba uma grande diversidade de formas de RNA, que se agrupam em RNA codificante e não codificante. O primeiro é o RNA mensageiro (mRNA) que tem como função servir de modelo para a síntese de proteínas na célula. Por sua vez, o RNA não codificante, constitui 98% do transcritoma e inclui não só o RNA de transferência (tRNA) e o RNA ribossomal (rRNA), essenciais na tradução, mas também os RNAs nucleolares (snoRNAs), Vault RNAs (vRNAs), PIWI-interacting RNAs (piRNAs) e ainda os pequenos RNAs (sRNAs), onde se incluem várias formas de RNAs de interferência (siRNAs) e microRNAs (miRNAs).
Os microRNAs são das mais pequenas moléculas de RNA não codificante, sendo constituídos apenas por 20-24 nucleótidos. Foram descobertos em 1993 em estudos no nemátode Caenorhabditis elegans, aquando da análise dos genes lin-14 e lin-4, envolvidos no desenvolvimento deste organismo modelo1. Nas plantas, o primeiro miRNA foi identificado em Arabidopsis thaliana (L.) Heynh.2 e até à data foram já identificadas mais de cerca de 8500 destas moléculas em mais de 70 espécies. A expressão dos miRNAs é altamente dependente do tipo de tecido e, por isso, os seus padrões e timings de expressão variam ao longo do desenvolvimento. Estão envolvidos no controlo da expressão genética, atuando pós- -transcricionalmente. Por exemplo, em mamíferos, a expressão de 60% dos genes codificantes de proteínas é regulada por miRNAs3. Muitos miRNAs pertencem a famílias cujas sequências são muito similares entre os membros, variando apenas num único nucleótido. Por sua vez, cada miRNA atua em vários mRNAs-alvo, sendo que nas plantas os miRNAs têm, em geral, menos genes-alvo que os animais. Além disso, cada mRNA é alvo da ação de diferentes miRNAs, resultando assim numa complexa rede de regulação4, 5.
Os miRNAs são codificados pelos genes MIRNAs (MIR), muitos dos quais bastante conservados no Reino Vegetal6. Transcritos pela RNA POLIMERASE II, os transcritos MIR de cadeia simples, dobram-se sobre si mesmos emparelhando e formando uma estrutura semelhante a um gancho de cabelo (harpin ou stem-loop), passando assim a designar-se miRNAs primários (pri-miRNAs)7. Nas plantas, os pri-miRNAs são clivados pela ação da RNase DICER-LIKE 1 (DCL1), daí resultando pequenas porções de RNA de cadeia dupla, os precursores dos miRNAs (pre-miRNAs). Estes sofrem depois metilação nas extremidades 3’ pela ação da RNA metiltransferase HUA ENHANCER 1 (HEN1), tornando-se maduros e, consequentemente, saem do núcleo através de uma proteína homóloga da exportina, designada HASTY8. Uma vez no citoplasma, ligam-se à proteína ARGONAUTE 1 (AGO1) e, em conjunto com outras proteínas, formam o complexo de silenciamento induzido por RNA (RISC, RNA-induced silencing complex), que identifica os mRNAs-alvo. Por complementaridade quase perfeita de bases nucleotídicas, na região codificante ou na região 5’ não traduzida (UTR) do mRNA, os miRNAs induzem silenciamento genético4. Este silenciamento ocorre nas células através de dois mecanismos: da repressão da tradução inibindo a síntese proteica ou, na sua maioria, da degradação do mRNA através da sua clivagem (FIGURA 1).
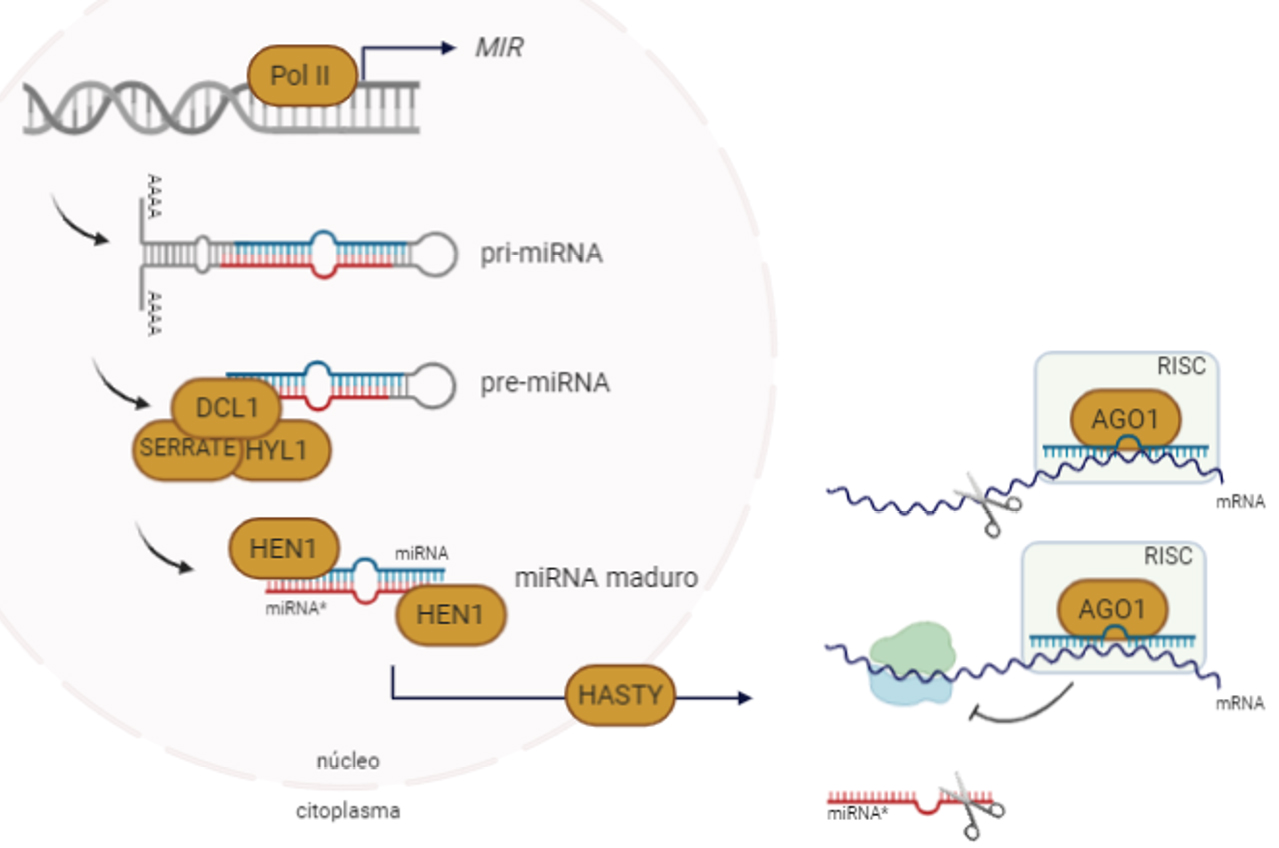
Provou-se, que o silenciamento genético pela ação de miRNAs é fundamental na regulação de muitos processos biológicos e não apenas como função de defesa das células contra agentes patogénicos como, por exemplo, os vírus. Através da modulação da expressão de genes, os miRNAs contribuem para a regulação da homeostase da célula, controlam processos biológicos e de desenvolvimento, como a diferenciação e manutenção da identidade e plasticidade de tecidos, e atuam ainda em respostas ao stress biótico e abiótico9. Os miRNAs exercem funções através do controlo da expressão temporal e espacial dos genes-alvo, principalmente os que têm uma função reguladora como os que codificam fatores de transcrição. Este tipo de interação miRNA−gene-alvo tem vindo a ser cada vez mais investigado e tem-se revelado determinante na compreensão de vários mecanismos moleculares que ocorrem na célula.
Nas plantas, a ação de miRNAs está já bem estudada em todo o processo do desenvolvimento da folha e sabe-se que controlam mecanismos desde a iniciação até à senescência, incluindo a diferenciação, polaridade, crescimento, forma e filotaxia10. Por exemplo, o miR319 está envolvido na morfogénese das folhas através da regulação das proteínas TCP. Também a determinação da diferenciação dorso-ventral, i.e., página superior ou inferior, é regulada pela ação do miR166 no gene PHB11, enquanto na raiz esta mesma interação miR- 166-PHB afeta o estabelecimento do padrão radial de desenvolvimento12. Já a identidade dos órgãos florais é controlada por AP2, regulado pelo miR17213.
Os miRNAs estão também envolvidos no controlo da transição da fase juvenil para a fase adulta nas plantas. No entanto, e apesar do crescente número de estudos sobre a importância dos miRNAs no ciclo de vida das plantas, o papel funcional destas moléculas no desenvolvimento embrionário não é ainda claro. Porém, estudos comprovam que múltiplos miRNAs são essenciais na morfogénese do embrião bem como no controlo da sua maturação. De facto, mutações nas proteínas envolvidas na biogénese de miRNAs resultam em alterações nos estádios primordiais do desenvolvimento do embrião e na incapacidade de indução da embriogénese somática, revelando assim a importância dos miRNAs na regulação do processo de embriogénese. Além disso, tem sido identificada uma expressão diferencial de vários miRNAs em culturas embriogénicas de diferentes gimnospérmicas, monocotiledóneas e dicotiledóneas, incluindo espécies economicamente importantes. Também muitas interações miRNAs−genes-alvo são determinantes durante este processo, principalmente aquelas relacionadas com o mecanismo de transdução de sinal da auxina14. Corroborando estes dados, estudos de embriogénese somática em Arabidopsis thaliana demonstraram que o miR393 controla a transição embriogénica de células somáticas através da regulação dos recetores de auxina TRANSPORT INHIBITOR1 e AUXIN F-BOX PROTEIN115, modificando a sensibilidade dos explantes ao tratamento de auxina. Também o miR396 controla a resposta embriogénica de células somáticas regulando os fatores de transcrição GROWTH-REGULATING FACTOR16 através de uma via relacionada com as auxinas. Já o miR167 atua através da regulação da expressão dos componentes de sinalização de auxina AUXIN RESPONSE FACTOR 6 e 817.
O estudo de miRNAs tem constituído, nos últimos anos um desafio para a comunidade científica, uma vez que as técnicas de biologia molecular comummente aplicadas não tinham a especificidade e sensibilidade necessária para a deteção de moléculas tão pequenas. Hoje, estão disponíveis técnicas que permitem a análise funcional de miRNAs de plantas, incluindo a sua análise in silico bem como a extração, deteção, quantificação, localização e análise da sua atividade no contexto celular18.
Em 2007, descobriu-se que os miRNAs também são alvo de regulação pós-transcripcional. Assim, Franco-Zorrilla e os seus colaboradores demonstraram, em A. thaliana, que o gene IPS1 tem complementaridade imperfeita com a sequência do miR399, o que leva ao seu sequestro, impossibilitando-o de degradar o mRNA do gene alvo (PHO2)19. Este tipo de mimetismo de alvo natural por parte do IPS1 conduziu ao desenvolvimento de miméticos sintéticos que permitem controlar a ação de outros miRNAs. Esta descoberta constitui, pois, uma ferramenta extremamente útil no estudo funcional de miRNAs, além de ter uma aplicação prática como, por exemplo, na modificação de características fenotípicas das plantas ou mesmo como agentes terapêuticos no tratamento de doenças humanas associadas a miRNAs, como é o caso da redução dos níveis de colesterol no plasma.
Além disso, nas plantas, os miRNAs podem ser introduzidos estavelmente no genoma, para seletivamente silenciarem um ou mais genes. Esta técnica tem sido aplicada na produção de plantas resistentes a infeções20 e tolerantes a stresses abióticos, tornando-as, por exemplo, resistentes a condições de salinidade21 ou de privação de nutrientes22. Por outro lado, tem-se revelado como uma solução para a eliminação de compostos indesejáveis em plantas alimentares, de que é exemplo a eliminação de alergénios do amendoim, e em plantas que se podem tornar comestíveis, como é o caso das sementes de algodão que contêm compostos tóxicos23.
Concluindo, o estudo dos miRNAs e da sua capacidade de modulação da expressão de genes tem-se revelado uma ferramenta molecular importante com forte impacto no melhoramento lhoramento de espécies agrícolas e em terapias médicas, esperando-se que nos próximos anos este tipo de moléculas possa vir a ter novas aplicações práticas na área da saúde e na agricultura.
Referências
- 1 LEE RC, et al. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14, Cell, 75, 843–854. 1993.
- 2 REINHART, B. J. et al., MicroRNAs in plants, Gene Dev, 16, 1616–1626. 2002.
- 3 FRIEDMAN, R. C. et al. Most mammalian mRNAs are conserved targets of microRNAs, Genome Res, 19, 92-105. 2009.
- 4 MILLAR, A. A. & WATERHOUSE, P. M., Plant and animal microRNAs: similarities and differences, Funct Integr Genomic, 5, 129-135. 2005.
- 5 PETER, M. E., Targeting of mRNAs by multiple miRNAs: the next step, Oncogene, 29, 2161-2164. 2010.
- 6 JONES-RHOADES, M. W., Conservation and divergence in plant microRNAs, Plant Mol Biol, 80, 3–16. 2012.
- 7 XIE, Z., Expression of Arabidopsis MIRNA genes, Plant Physiol, 138, 2145–2154. 2005.
- 8 PARK, M.Y. et al., Nuclear processing and export of microRNAs in Arabidopsis, P Natl Acad Sci USA, 102, 3691–3696. 2005.
- 9 SUN, G., MicroRNAs and their diverse functions in plants, Plant Mol Biol, 80, 17–36. 2012.
- 10 PULIDO, A. & LAUFS, P., Co-ordination of developmental processes by small RNAs during leaf development, J Exp Bot, 61, 1277–1291. 2010.
- 11 KIDNER, C. & MARTIENSSEN, R., Spatially restricted microRNA directs leaf polarity through ARGONAUTE1, Nature, 428, 81–84. 2004.
- 12 CARLSBECKER, A. et al., Cell signalling by microRNA165/6 directs gene dose-dependent root cell fate, Nature, 465, 316-321. 2010.
- 13 MILO, JA. & HAJIME, S., Regulation of flowering time and floral organ identity by a microRNA and its APETALA2-like target genes, The Plant Cell, 15, 2730-2741. 2003.
-
14 WÓJCIKOWSKA, B.
- 15 WÓJCIK, A. M. & GAJ, M. D., miR393 contributes to the embryogenic transition induced in vitro in Arabidopsis via the modification of the tissue sensitivity to auxin treatment, Planta, 244, 231-243. 2016.
- 16 SZCZYGIE?-SOMMER, A. & GAJ, M. D., The miR396-GRF regulatory module controls the embryogenic response in Arabidopsis via an auxin-related pathway, Int J Mol Sci, 20, 5221. 2019.
- 17 SZYRAJEW, K. et al. MicroRNAs are intensively regulated during induction of somatic embryogenesis in Arabidopsis, Front Plant Sci, 8, 18. 2017.
- 18 WÓJCIK, A. M., Research tools for the functional genomics of plant miRNAs during zygotic and somatic embryogenesis, Int J Mol Sci, 21, 4969. 2020.
- 19 FRANCO-ZORRILLA, J. M. et al. Target mimicry provides a new mechanism for regulation of microRNA activity, Nat Genet, 39, 8, 1033–1037. 2007.
- 20 HUANG, G., et al., Engineering broad root-knot resistance in transgenic plants by RNAi silencing of a conserved and essential root-knot nematode parasitism gene, P Natl Acad Sci USA, 103, 14302–14306. 2006.
- 21 ZHOU, M. et al. Constitutive expression of a miR319 gene alters plant development and enhances salt and drought tolerance in transgenic creeping bentgrass, Plant Physiol, 161, 1375-1391. 2013.
- 22 YUAN, S. et al. Constitutive expression of rice microRNA528 alters plant development and enhances tolerance to salinity stress and nitrogen starvation in creeping bentgrass, Plant Physiol, 169, 576-593. 2015.
- 23 SUNILKUMAR, G. et al., Engineering cottonseed for use in human nutrition by tissue-specific reduction of toxic gossypol, P Natl Acad Sci USA, 103, 18054-18059. 2006.
Este artigo já foi visualizado 9692 vezes.